Endosporos estão adormecidos, não-reprodutivos e enzimaticamente inertes de células vegetativas bacterianas . As bactérias produtoras de endosporos são mais convencionalmente denominadas “bactérias esporulantes”. As bactérias esporulantes são derivadas do filo Firmicute e consistem principalmente em bactérias Gram positivas . Estas bactérias produtoras de endosporo pertencem ao género Bacillus e Clostridium . Um endosporo é uma forma reduzida de uma bactéria. É constituído por um núcleo contendo apenas os elementos essenciais necessários para o início do crescimento bacteriano. O núcleo é rodeado por um revestimento resistente e altamente impermeável que é composto por várias camadas. Os endosporos servem para proteger a bactéria de condições ambientais nocivas, reduzindo-a a um estado dessecado, criptobiótico e altamente defensivo que transmite resistência a muitas condições ambientais que de outra forma prejudicariam e matariam a forma vegetativa da bactéria. Estas condições ambientais incluem temperaturas extremas, radiação, níveis de pH extremos, pressões extremas e agentes químicos nocivos. A dormência metabólica e a capacidade protectora dos endosporos foram demonstradas pelo isolamento e renascimento bem sucedido dos endosporos Bacillus sp. de 25 a 40 milhões de anos de idade âmbar . Assim, a esporulação provou ser um mecanismo vital de sobrevivência bacteriana.
br>>>h2>O Ciclo de Vida das Bactérias Esportivas
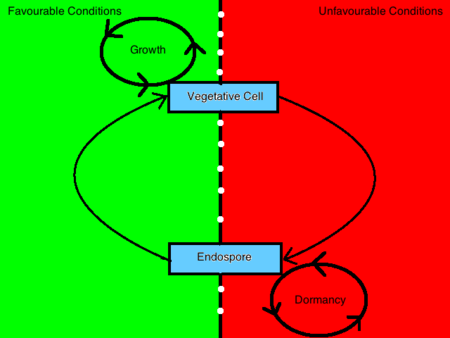
As bactérias esporulantes podem assumir duas morfologias distintas, uma sendo um endosporo e a outra sendo uma célula vegetativa. A decisão de se conformar a qualquer uma das morfologias é determinada por estímulos ambientais que transmitem informação para o interior da célula através de um processo mediado por um sistema de transdução de sinal de fósforo. As células bacterianas irão esporular em resposta a condições de crescimento debilitantes . Em contraste, uma bactéria vegetará na presença de germinantes, nutrientes e outras condições ambientais favoráveis . Quando uma bactéria sente uma mudança desfavorável nas condições ambientais de tal forma que o crescimento já não é alcançável, transmitirá esta informação para o interior da célula através da interacção de um sensor cinase e de uma proteína reguladora e induzirá a regulação dos produtos genéticos esporulantes . Esta célula, por sua vez, sofre uma transformação que a tornará resistente a essas condições ambientais e lhe permitirá permanecer em estado dormente até que as condições favoráveis sejam restauradas.
Mecanismo da esporulação
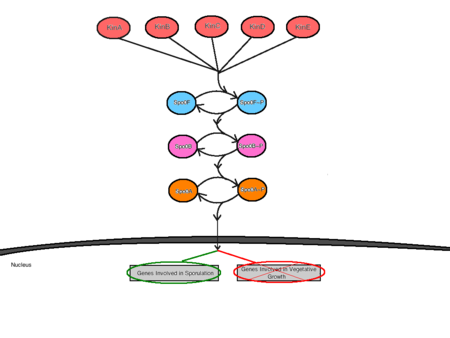
A iniciação da esporulação é mediada por pelo menos cinco kinases sensoras que se alimentam da cascata de transdução do sinal de fósforo da esporulação . Estas kinases sensoriais são denominadas KinA, KinB, KinC, KinD e KinE, e são cada uma específica para diferentes estímulos ambientais . A existência de várias quinases sensoriais permite que múltiplos sinais ambientais influenciem o início da esporulação . Em resposta à interacção específica com estímulos ambientais desfavoráveis, as cinases sensoriais autofosforam um resíduo conservado de histidina através de uma reacção dependente de ATP . As diferentes quinases sensoriais alimentam os seus respectivos sinais na via do fósforo através de uma única proteína chamada Spo0F . A partir daqui, Spo0F~P transfere o grupo fosfato para Spo0B e finalmente, Spo0B~P fosforilatos Spo0A . Spo0B actua como um mediador entre Spo0F e Spo0A, fornecendo os sinais desde o ponto de entrada na célula até ao ponto de regulação. A Spo0A actua como uma proteína reguladora de ligação ao ADN, regulando positivamente os genes necessários para a esporulação e negativamente os genes envolvidos no crescimento vegetativo .
Estrutura endospore
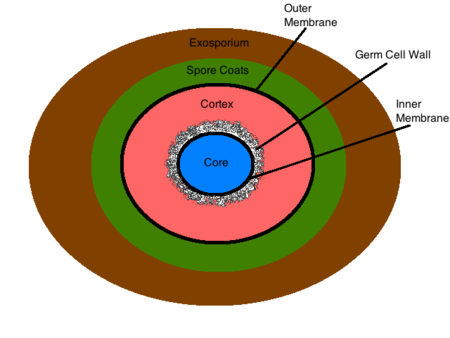
O exosporium é a estrutura mais externa do esporo, constituída principalmente por proteínas, lípidos e hidratos de carbono . Esta composição confere ao exosporium a sua característica hidrofóbica inerente que o torna insolúvel e contribui para a patogenicidade dos esporos . Esta estrutura não está presente em todas as bactérias esporulantes . A camada de esporos encontra-se debaixo do exosporio (se presente) e é feita de finas camadas proteicas que são compostas de proteínas da camada que estão fortemente ligadas através de ligações dissulfeto. Esta estrutura serve como barreira inicial de permeabilidade, peneirando as moléculas maiores e permitindo a permeação de moléculas mais pequenas, tais como germinantes . A membrana exterior reside sob a camada de esporos e a sua função ainda não é conhecida . O córtex encontra-se abaixo da membrana exterior e é uma estrutura peptidoglicano que difere do peptidoglicano vegetativo pela sua falta de ácidos teicóicos e baixos níveis de ligação cruzada . A parede celular germinativa encontra-se abaixo do córtex e é também composta de peptidoglicano . As diferenças estruturais na parede celular germinal e no córtex permitem a degradação selectiva do córtex após a germinação e facilitam a transformação da parede celular germinal na parede celular bacteriana de uma célula vegetativa . Por baixo da parede da célula germinal encontra-se a membrana interna. Esta estrutura é semelhante em composição à membrana citoplasmática de uma célula vegetativa, mas difere muito na sua fluidez e permeabilidade. A mobilidade reduzida dos lípidos da membrana torna a estrutura altamente impermeável a todas as moléculas incluindo a água, impedindo assim a difusão da maioria das moléculas para o núcleo . A membrana interna contém receptores germinantes de superfície que ligam os germinantes e iniciam a germinação e o crescimento vegetativo . O núcleo contém o ADN bacteriano, RNA, ribossomas e a maioria das enzimas essenciais . O seu estado desidratado contribui para a dormência enzimática e resistência ao calor do esporo . Dentro do núcleo, o DNA bacteriano está ligado a pequenas proteínas de esporos solúveis em ácido (SASPs) que protegem fisicamente o DNA de substâncias químicas e enzimas nocivas . As condições dentro do núcleo do esporo contribuem largamente para a sua resistência a condições ambientais desfavoráveis, e são discutidas em detalhe abaixo.
Resistência dos Endosporos
Resistência ao calor húmido é uma característica notável dos esporos quando suspensos num meio aquoso. Geralmente, a resistência ao calor húmido permite que os esporos sejam resistentes a temperaturas 40 graus celsius superiores às suas respectivas formas vegetativas. O principal factor determinante da resistência ao calor húmido é o teor de água no núcleo. Os esporos apresentam uma relação inversamente proporcional entre o conteúdo de água e a resistência ao calor húmido, pelo que os esporos com maior resistência ao calor húmido têm diminuído o conteúdo do núcleo aquoso . Além disso, os esporos que se formam a temperaturas mais elevadas terão naturalmente um menor teor de água no núcleo e, por conseguinte, uma maior resistência ao calor húmido. O calor seco causa danos significativos no DNA de uma célula vegetativa . Como resultado, os SASPs desempenham um papel importante na resistência ao calor seco dos esporos. Ao ligar e proteger fisicamente o ADN, os SASP previnem a sua quebra a temperaturas elevadas .
Resistência química
Muitas substâncias químicas nocivas matam bactérias através de mecanismos que danificam o ADN . No entanto, os esporos desenvolveram uma série de mecanismos que conferem resistência química. A camada de esporos é importante na resistência a muitas substâncias químicas oxidantes tais como dióxido de cloro, hipoclorito, ozono e peroxinitrito . Funciona reagindo com estes agentes químicos e desintoxicando-os antes de estes passarem pela camada de esporos . Além disso, a permeabilidade extremamente baixa da membrana interna impede tanto as moléculas hidrofóbicas como as hidrofílicas de entrar no núcleo . Os SASP também protegem o ADN ligando-se a ele e protegendo-o de agentes químicos nocivos que entram no núcleo .
Resistência à radiação UV
A radiação UV danifica o ADN de uma célula e induz mutações. A saturação do ADN de um endosporo com SASP protege o ADN desta ameaça nociva .
(1)Cano, Raul J., e Monica K. Borucki. “Renascimento e identificação de esporos bacterianos em âmbar dominicano de 25 a 40 milhões de anos de idade”. Ciência 268.5213 (n.d.) (2012): 1060-064. Web.
(2)Leggett, M.J., G. McDonnell, S.P. Denyer, P. Setlow, e J.-Y. Maillard. “Estruturas de esporos bacterianos e o seu papel protector na resistência aos biocidas”. Journal of Applied Microbiology 113 (2012): 485-98. Web.
(3)Stephenson, Keith, e James A. Hoch. “Evolução da Sinalização no Fósforo de Esporulação”. Microbiologia Molecular 2 (2002): 2967-304. Web.
(4) Piggot, Patrick J., e David W. Hilbert. “Sporulation of Bacillus Subtilis”. Opinião actual em Microbiolgy 7 (2004): 579-86. Web.
(5)http://www.eoearth.org/article/Bacteria?topic=49480#gen0
(6)Varghuese, Kottayil I., Haiyan Zhao, Vidya Harini Veldore, e James Zapf. “Sporulation Phosphorelay Proteins and Their Complexes” (Proteínas de Fósforo Esportivo e seus Complexos): Caracterização Cristalográfica”. Métodos em Enzimologia 422 (2007): 102-22. Web.
(7) Hilbert, David W., e Patrick J. Piggot. “Compartimentação da Expressão do Género durante a Formação de Esporos de Bacillus Subtilis”. Microbiology and Molecular Biology Reviews (2004): 234-62. Web.
(8)Stragier, Patrick, e Richard Losick. “Molecular Genetics of Sporulation in Bacillus Subtilis”. Revisões Anuais 30 (1996): 297-341. Web.
(9)Setlow, Peter. “Germinação de esporos”. Current Opinion in Microbiolgy 6 (2003): 550-56. Web.
(10)Setlow, Peter. “Esporos de Bacillus Subtilis”: A sua Resistência e Matança por Radiação, Calor e Químicos”. Journal of Applied Microbiology 101 (2006): 514-25. Web.
(11) Nicholson, Wayne L., Nobuo Munakata, Gerda Horneck, Henry J. Melosh, e Peter Setlow. “Resistência de Bacillus Endospores aos Ambientes Extremos Terrestre e Extraterrestre”. Microbiologia e Biologia Molecular Revisões 64 (2000): 548-72. Web.