Le endospore sono forme dormienti, non riproduttive ed enzimaticamente inerti di cellule vegetative batteriche. I batteri che producono endospore sono più convenzionalmente chiamati “batteri sporulanti”. I batteri sporulanti derivano dal phylum Firmicute e consistono principalmente in batteri Gram positivi. Questi batteri endosporei appartengono ai generi Bacillus e Clostridium. Un’endospora è una forma ridotta di un batterio. È composta da un nucleo che contiene solo lo stretto necessario per l’inizio della crescita batterica. Il nucleo è circondato da un rivestimento duro e altamente impermeabile composto da diversi strati. Le endospore servono a proteggere il batterio da condizioni ambientali dannose riducendolo in uno stato essiccato, criptobiotico e altamente difensivo che trasmette resistenza a molte condizioni ambientali che altrimenti danneggerebbero e ucciderebbero la forma vegetativa del batterio. Queste condizioni ambientali includono temperature estreme, radiazioni, livelli estremi di pH, pressioni estreme e agenti chimici dannosi. La dormienza metabolica e la capacità protettiva delle endospore sono state dimostrate dall’isolamento e dalla rinascita di endospore di Bacillus sp. da un’ambra di 25-40 milioni di anni. Così, la sporulazione ha dimostrato di essere un meccanismo vitale di sopravvivenza batterica.
Il ciclo di vita dei batteri sporulanti
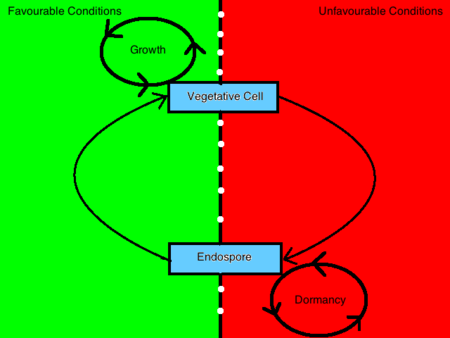
I batteri sporizzanti possono assumere due morfologie distinte, una è un’endospora e l’altra è una cellula vegetativa. La decisione di conformarsi all’una o all’altra morfologia è determinata da stimoli ambientali che trasmettono informazioni all’interno della cellula attraverso un processo mediato da un sistema di trasduzione del segnale fosforico. Le cellule batteriche sporuleranno in risposta a condizioni di crescita debilitanti. Al contrario, un batterio vegeterà in presenza di germinanti, nutrienti e altre condizioni ambientali favorevoli. Quando un batterio percepisce un cambiamento sfavorevole nelle condizioni ambientali tale che la crescita non è più raggiungibile, trasmetterà questa informazione all’interno della cellula attraverso l’interazione di un sensore chinasi e una proteina regolatrice e indurrà l’up-regolazione dei prodotti genici sporulanti. Questa cellula, a sua volta, subisce una trasformazione che la renderà resistente a quelle condizioni ambientali e le permetterà di rimanere in uno stato dormiente fino al ripristino delle condizioni favorevoli.
Meccanismo della sporulazione
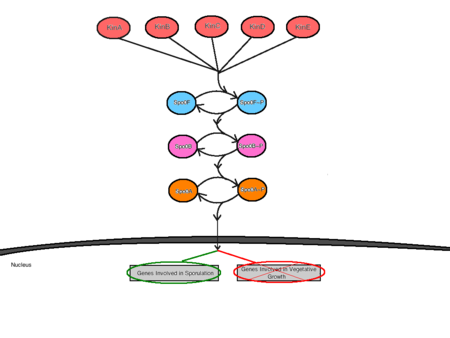
L’inizio della sporulazione è mediato da almeno cinque sensori chinasi che alimentano la cascata di trasduzione del segnale di fosforeggiamento della sporulazione. Queste chinasi sensore sono denominate KinA, KinB, KinC, KinD e KinE, e sono ciascuna specifica per diversi stimoli ambientali. Avere diverse chinasi sensore permette per più segnali ambientali per influenzare l’inizio della sporulazione. In risposta all’interazione specifica con stimoli ambientali sfavorevoli, le chinasi sensore si autofosforilerebbero un residuo di istidina conservato mediante una reazione ATP-dipendente. Le diverse chinasi sensore alimentano i loro rispettivi segnali nella via di fosforilazione attraverso una singola proteina chiamata Spo0F. Da qui, Spo0F~P trasferisce il gruppo fosfato a Spo0B e infine, Spo0B~P fosforila Spo0A . Spo0B agisce come mediatore tra Spo0F e Spo0A consegnando i segnali dal punto di ingresso nella cellula al punto di regolazione. Spo0A agisce come una proteina regolatrice legata al DNA, regolando positivamente i geni necessari alla sporulazione e negativamente i geni coinvolti nella crescita vegetativa.
Struttura dell’endospora
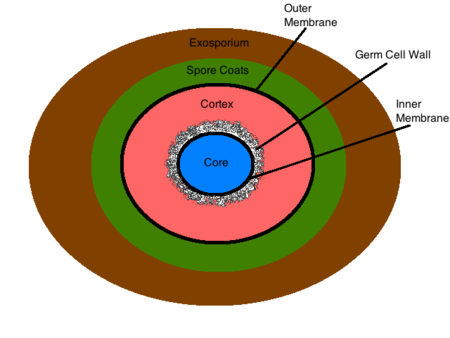
L’esosporio è la struttura più esterna della spora, composta principalmente da proteine, lipidi e carboidrati. Questa composizione conferisce all’esosporio la sua caratteristica idrofoba intrinseca che lo rende insolubile e contribuisce alla patogenicità delle spore. Questa struttura non è presente in tutti i batteri sporulanti. Il mantello della spora si trova sotto l’esosporio (se presente) ed è costituito da sottili strati proteici che sono composti da proteine del mantello che sono pesantemente legati tra loro tramite legami disolfuro. Questa struttura serve come barriera di permeabilità iniziale, setacciando le molecole più grandi e permettendo la permeazione di molecole più piccole come i germogli. La membrana esterna risiede sotto il mantello della spora e la sua funzione non è ancora nota. La corteccia si trova sotto la membrana esterna ed è una struttura di peptidoglicano che differisce dal peptidoglicano vegetativo per la sua mancanza di acidi teicoici e bassi livelli di legame incrociato. La parete delle cellule germinali si trova sotto la corteccia ed è anche composta da peptidoglicano. Le differenze strutturali nella parete cellulare del germe e nella corteccia permettono la degradazione selettiva della corteccia al momento della germinazione e facilitano la trasformazione della parete cellulare del germe nella parete cellulare batterica di una cellula vegetativa. Sotto la parete cellulare del germe si trova la membrana interna. Questa struttura è simile nella composizione alla membrana citoplasmatica di una cellula vegetativa, ma differisce notevolmente nella sua fluidità e permeabilità. La ridotta mobilità dei lipidi di membrana rende la struttura altamente impermeabile a tutte le molecole, compresa l’acqua, impedendo così la diffusione della maggior parte delle molecole nel nucleo. La membrana interna contiene recettori germinativi di superficie che legano i germinativi e avviano la germinazione e la crescita vegetativa. Il nucleo contiene il DNA batterico, l’RNA, i ribosomi e la maggior parte degli enzimi essenziali. Il suo stato disidratato contribuisce alla dormienza enzimatica e alla resistenza al calore della spora. All’interno del nucleo il DNA batterico è legato a piccole proteine acido-solubili della spora (SASPs) che proteggono fisicamente il DNA da sostanze chimiche ed enzimi nocivi. Le condizioni all’interno del nucleo della spora contribuiscono in gran parte alla sua resistenza a condizioni ambientali sfavorevoli, e sono discusse in dettaglio di seguito.
Resistenza delle endospore
La resistenza al calore umido è una caratteristica notevole delle spore quando sono sospese in un mezzo acquoso. Generalmente, la resistenza al calore umido permette alle spore di essere resistenti a temperature di 40 gradi celsius superiori alla loro rispettiva forma vegetativa. Il principale determinante della resistenza al calore umido è il contenuto di acqua del nucleo. Le spore presentano una relazione inversamente proporzionale tra il contenuto di acqua e la resistenza al calore umido e quindi, le spore con una maggiore resistenza al calore umido hanno un contenuto di acqua del nucleo diminuito. Inoltre, le spore che si formano a temperature più elevate avranno naturalmente un contenuto d’acqua del nucleo più basso, e quindi, hanno una maggiore resistenza al calore umido. Il calore secco provoca danni significativi al DNA in una cellula vegetativa. Di conseguenza, le SASP svolgono un ruolo importante nella resistenza al calore secco delle spore. Legandosi al DNA e proteggendolo fisicamente, le SASP impediscono la sua rottura a temperature elevate.
Resistenza chimica
Molte sostanze chimiche nocive uccidono i batteri attraverso meccanismi che danneggiano il DNA. Tuttavia, le spore hanno evoluto una serie di meccanismi che conferiscono resistenza chimica. Il mantello della spora è importante nella resistenza a molte sostanze chimiche ossidanti come il biossido di cloro, l’ipoclorito, l’ozono e il perossinitrito. Funziona reagendo con questi agenti chimici e disintossicandoli prima che passino attraverso il mantello della spora. Inoltre, la permeabilità estremamente bassa della membrana interna impedisce alle molecole idrofobiche e idrofile di entrare nel nucleo. Le SASP proteggono anche il DNA legandosi ad esso e schermandolo dagli agenti chimici nocivi che entrano nel nucleo.
Resistenza alle radiazioni UV
La radiazione UV danneggia il DNA di una cellula e induce mutazioni. La saturazione del DNA di un’endospora con i SASP protegge il DNA da questa minaccia dannosa.
(1)Cano, Raul J., e Monica K. Borucki. “Rinvenimento e identificazione di spore batteriche in un’ambra dominicana vecchia da 25 a 40 milioni di anni”. Scienza 268.5213 (n.d.) (2012): 1060-064. Web.
(2)Leggett, M.J., G. McDonnell, S.P. Denyer, P. Setlow, and J.-Y. Maillard. “Strutture delle spore batteriche e il loro ruolo protettivo nella resistenza ai biocidi”. Journal of Applied Microbiology 113 (2012): 485-98. Web.
(3)Stephenson, Keith, e James A. Hoch. “Evoluzione della segnalazione nel fosforello di sporulazione”. Molecular Microbiology 2 (2002): 2967-304. Web.
(4) Piggot, Patrick J., e David W. Hilbert. “Sporulazione di Bacillus Subtilis”. Current Opinion in Microbiolgy 7 (2004): 579-86. Web.
(5)http://www.eoearth.org/article/Bacteria?topic=49480#gen0
(6)Varghuese, Kottayil I., Haiyan Zhao, Vidya Harini Veldore, and James Zapf. “Proteine fosforiliche di sporulazione e i loro complessi: Caratterizzazione cristallografica”. Metodi in Enzimologia 422 (2007): 102-22. Web.
(7) Hilbert, David W., and Patrick J. Piggot. “Compartmentalizzazione dell’espressione genica durante la formazione delle spore di Bacillus Subtilis”. Microbiology and Molecular Biology Reviews (2004): 234-62. Web.
(8)Stragier, Patrick, e Richard Losick. “Genetica molecolare della sporulazione in Bacillus Subtilis”. Annual Reviews 30 (1996): 297-341. Web.
(9)Setlow, Peter. “Germinazione delle spore”. Current Opinion in Microbiolgy 6 (2003): 550-56. Web.
(10) Setlow, Peter. “Spore di Bacillus Subtilis: La loro resistenza e l’uccisione da parte di radiazioni, calore e sostanze chimiche”. Journal of Applied Microbiology 101 (2006): 514-25. Web.
(11) Nicholson, Wayne L., Nobuo Munakata, Gerda Horneck, Henry J. Melosh, e Peter Setlow. “Resistenza delle endospore di Bacillus agli ambienti estremi terrestri ed extraterrestri”. Microbiology and Molecular Biology Reviews 64 (2000): 548-72. Web.