h5>Dale Vogt, Helen A. Swartz e John Massey
Department of Animal Sciencesp>Vários esquemas de acasalamento de animais são classificados em duas grandes categorias – consanguinidade e surto. A classificação depende da proximidade da relação biológica entre companheiros. Dentro de cada categoria, existe uma grande variação na intensidade desta relação. Uma linha muito fina separa as duas categorias. O acasalamento de animais estreitamente relacionados (por exemplo, progenitor e descendência, irmão e irmã completos ou meio irmão e irmã) é consanguinidade. Com animais menos relacionados (primos em primeiro lugar, primos em segundo grau), as pessoas discordam sobre onde traçar a linha entre consanguinidade e surto.
Tecnicamente, a consanguinidade é definida como o acasalamento de animais mais estreitamente relacionados do que a relação média dentro da raça ou população em questão. Os acasalamentos entre animais menos próximos do que este, então, constituiriam um surto. Estes dois sistemas de acasalamento, com intensidades variáveis em cada um, são descritos no Quadro 1. Os acasalamentos indicados dentro da categoria de consanguinidade são auto-explicativos; aqueles dentro da categoria de surto são definidos no glossário.
Quadro 1
Graus de consanguinidade e de surto organizados de acordo com a relação biológica entre os acasalamentos indicados. (Ao ler de cima para baixo, a relação biológica entre os companheiros diminui constantemente.)
| Inbreeding or outbreeding | Coeficiente da relação entre companheiros | |
|---|---|---|
| Inbreeding | 50 por cento | Pai × descendência; irmãos completos |
| 25 por cento | Half-sibs; duplos primos em primeiro grau; tia × sobrinho; tio × sobrinha | |
| 12-1/2 por cento | Primeiro primo | |
| Consanguinidade | 6-1/4 por cento | Segundo primo |
| Consanguinidade | ? | Linebreeding1 |
| 0 por cento | acasalamento aleatório dentro da raça ou população2 | |
| 0 por cento | Eliminar | |
| Sangue | 0 por cento | |
| 0 por cento | Travessia de espécies | |
| Outbreeding | 0 por cento | cruzamento de géneros |
1Num programa de reprodução linfática, o coeficiente de relação entre companheiros é geralmente baixo; no entanto, pode ser bastante variável.
2O acasalamento aleatório dentro de uma raça ou população significa que os acasalamentos são escolhidos por acaso. Deve entender-se que sob esta circunstância é possível que ocorra consanguinidade ou surto.
Relações biológicas entre animais
Os indivíduos são considerados como biologicamente relacionados quando têm um ou mais antepassados comuns. Para efeitos práticos, se dois indivíduos não tiverem um antepassado comum nas últimas cinco ou seis gerações, são considerados não relacionados.
As relações biológicas são importantes na criação de animais porque quanto mais próxima a relação, maior a percentagem de genes semelhantes que os dois indivíduos carregam. A proximidade da relação é determinada por três factores:
- Quanto longe nos pedigrees dos dois animais o antepassado comum aparece
- Quantos antepassados comuns eles têm
- Quantas vezes os antepassados comuns aparecem. É também influenciado por qualquer consanguinidade do antepassado ou antepassados comuns
Medição do grau de relação biológica
O coeficiente de relação é um valor numérico único que considera todos os factores acima mencionados. É uma medida do grau de semelhança entre os genótipos (constituições genéticas) dos dois animais. É estimado pela expressão:
RBC = sigma ÷ Raiz quadrada de (1 + FB)(1 + FC)
Equação 1 onde:
RBC = o coeficiente de relação entre os animais B e C que queremos medir.
sigma = o símbolo grego que significa “adicionar”
(1/2) = a fracção de material genético de um indivíduo que é transmitida à sua descendência. É utilizado no cálculo do coeficiente de relações porque representa a probabilidade de, em qualquer geração, um gene idêntico de um dado par de genes ser transmitido para cada uma de duas descendência em particular. É também a probabilidade de um gene diferente de um dado par de genes ser transmitido aos dois descendentes.
n = o número de gerações entre o animal B e o antepassado comum.
n’ = o número de gerações entre o animal C e o antepassado comum
FA, FB, FC = coeficientes de consanguinidade do antepassado comum e dos animais B e C, respectivamente.
Se nenhum dos animais for consanguíneo, o coeficiente de relação é estimado como:
RBC = sigma
Equação 2
O uso desta expressão pode ser demonstrado com o pedigree completo da amostra de sexos e o diagrama de seta (Figura 1). Neste exemplo, suponha-se que nem o garanhão nem a barragem são consanguíneos. O diagrama de seta à direita mostra caminhos de fluxo de genes de cada um dos antepassados comuns (D e E) aos animais cujo coeficiente de relação estamos a medir (B e C).
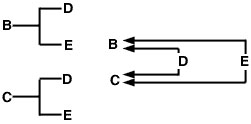
br>>igure 1
O problema agora é traçar todos os caminhos possíveis do animal B ao animal C que passam por um antepassado comum. Neste caso, existem dois desses caminhos (Figura 2).
Figure 2
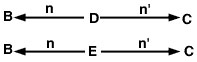 Duas vias
Duas vias
Equação 3
p>Desde que não assumimos a consanguinidade neste exemplo, o coeficiente de relação entre os animais B e C é estimado como:
p>RBC = sigma = (1/2)1 + 1 + (1/2)1 + 1 = (1/2)2 + (1/2)2 = 0.50
Utilidade do coeficiente de informação da relação
Um produtor de gado encontraria a informação do coeficiente de informação da relação valiosa em várias situações. Ele pode, por exemplo, querer vender um animal relacionado com um que anteriormente era vendido por um preço elevado. Quanto mais elevado for o coeficiente de relação entre os dois, melhor será a sua utilização como ponto de venda. Ou, pode querer comprar um dos dois touros relacionados e um pode custar mais do que deseja pagar. Se o coeficiente de relação entre os dois touros for elevado, ele pode estar tão bem com o touro de preço mais baixo como estaria com o mais caro.
Uma utilização prática do coeficiente de relação é estimar o valor de desempenho de um animal não testado. Para estimar o valor, devemos conhecer o valor do desempenho de um animal relacionado, o coeficiente de relação entre os animais testados e não testados, e o valor médio do desempenho da raça, manada, ou grupo a que pertencem os animais testados e não testados.
Como exemplo, considere um manada com um ganho médio diário de 2,25 libras (ADG) por dia. Assumir ainda que um reprodutor deste rebanho tinha um feedlot ADG de 3,50 libras por dia, enquanto um meio-irmão mais novo ainda não foi avaliado para feedlot ADG. Assumindo que não houve consanguinidade, o coeficiente de relação entre meios-irmãos é de 0,25. A melhor estimativa do feedlot ADG do animal não testado é que se desviará da média do efectivo em 25 por cento, na medida em que o valor do desempenho do meio-irmão testado. Utilizando estes valores, o valor mais provável do feedlot ADG do animal não testado é 2,25 + (0,25) × (3,50 – 2,25) ou 2,56 libras por dia.
Medição do grau de consanguinidade
Quando calculamos um coeficiente de consanguinidade, estamos a tentar medir a percentagem provável de redução na frequência de emparelhamento de genes diferentes (redução na heterozigosidade). Esta redução é relativa a uma população de base. A população de base é geralmente a raça em questão numa data em que os pedigrees são rastreados. Assume-se que os animais desta população de base não são criados. Isto não significa que estes animais da população de base tenham genes diferentes em cada par. Não há forma de sabermos quantos dos seus pares de genes consistiam em genes semelhantes ou dissimilares. O coeficiente de consanguinidade que é calculado é simplesmente relativo a essa base e reflecte a provável redução percentual em muitos pares de genes díspares que os animais da população base média tinham.
A expressão geral para determinar o coeficiente de consanguinidade é:
FX = sigma
Equação 4 onde:
FX = o coeficiente de consanguinidade do animal X.
sigma = o símbolo grego que significa “adicionar”.
(1/2) = a fracção do material genético de um indivíduo que é transmitido à sua descendência. É utilizado no cálculo do coeficiente de relações porque representa a probabilidade de, em qualquer geração, um gene idêntico de um dado par de genes ser transmitido para cada uma de duas descendência em particular. É também a probabilidade de um gene diferente de um dado par de genes ser transmitido aos dois descendentes.
n = o número de gerações entre o animal B e o antepassado comum.
n’ = o número de gerações entre o animal C e o antepassado comum.
h5>+1 = é adicionado a n e n’ para contabilizar a geração adicional entre o animal × e os seus progenitores.
FA = o coeficiente de consanguinidade do ascendente comum.
Se nenhum dos progenitores for consanguíneo, mas se forem parentes, o coeficiente de consanguinidade da sua prole é metade do seu coeficiente de parentesco: 1/2 RBC Isto pode ser demonstrado utilizando um acasalamento de sexos completo para facilitar a comparação com o coeficiente de relação de sexos completo calculado anteriormente. Neste caso, o pedigree para o animal X e o diagrama de seta será o seguinte:
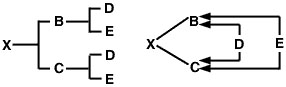
Figure 3
O problema agora é traçar todos os caminhos possíveis desde o reprodutor (B) até à barragem (C) através de cada antepassado comum. Tal como com o problema do coeficiente de relação, existem dois desses caminhos:
![]() Figura 4
Figura 4
P>Desde que assumimos que nenhum dos progenitores é consanguíneo, o coeficiente de consanguinidade do animal × é estimado como:
FX = sigma = (1/2)1 + 1 + 1 + (1/2)1 + 1 + 1 = (1/2)3 + (1/2)3 = 0.25
Equação 5
É metade do coeficiente de relação entre sexos inteiros quando não há consanguinidade.
Consequências genéticas da consanguinidade
A consequência genética básica da consanguinidade é promover o que é tecnicamente conhecido como homozigocidade. Isto significa que há um aumento na frequência de emparelhamento de genes semelhantes. Acompanhando este aumento, deve haver uma diminuição na frequência de emparelhamento de genes diferentes. A isto chama-se uma diminuição da heterozigosidade. Estes eventos simultâneos são as razões subjacentes aos efeitos gerais no desempenho que observamos com a consanguinidade.
Razões para a consanguinidade
Desenvolvimento de linhas consanguíneas altamente produtivas de gado doméstico é possível. Até à data, porém, tais tentativas têm tido pouco sucesso aparente. Embora ocasionalmente sejam produzidos animais de alto rendimento, a consanguinidade resulta geralmente numa redução global do rendimento. Esta redução manifesta-se de muitas maneiras. Os efeitos mais óbvios da consanguinidade são uma menor eficiência reprodutiva, incluindo taxas de mortalidade mais elevadas, taxas de crescimento mais baixas e uma maior frequência de anomalias hereditárias. Isto tem sido demonstrado por numerosos estudos com bovinos, equinos, ovinos, suínos e animais de laboratório.
A extensão desta diminuição no desempenho, em geral, é proporcional ao grau de consanguinidade. Quanto maior for o grau de consanguinidade, maior é a redução do desempenho. A redução real do desempenho não é a mesma em todas as espécies ou em todas as características. Algumas características (como a qualidade da carne) dificilmente são influenciadas pela consanguinidade; outras (como a eficiência reprodutiva) são grandemente influenciadas pela consanguinidade. Não podemos, portanto, fazer uma afirmação generalizada sobre a quantidade de redução no “desempenho” que resultaria de uma quantidade específica de consanguinidade e esperar que seja aplicável numa grande variedade de situações.
É possível, contudo, prever a extensão do efeito da consanguinidade sobre características específicas. Tais previsões baseiam-se em resultados efectivamente obtidos em condições experimentais em que vários níveis de consanguinidade consanguínea tinham sido atingidos. Em pesquisas com suínos realizadas no Laboratório Regional de Reprodução Suína do Midwest, Dickerson e outros (1954) apontam que para cada 10 por cento de aumento na consanguinidade (dos suínos da ninhada), há uma diminuição de 0,20, 0,35, 0,38, e 0,44 porcos por ninhada ao nascimento, 21 dias, 56 dias, e 154 dias, respectivamente. Podemos utilizar estes números para fornecer estimativas das diminuições esperadas no tamanho da ninhada (em idades comparáveis) noutros rebanhos de suínos.
Os estudos de consanguinidade sugerem que cada aumento sucessivo da consanguinidade resulta numa diminuição proporcional do desempenho. As estimativas de aumentos médios na percentagem de consanguinidade dentro de um efectivo fechado podem ser feitas com a expressão:
Avg. aumento na consanguinidade = (Número de machos + Número de fêmeas) ÷ (8)(Número de machos)(Número de fêmeas)
Num rebanho fechado de gado em que 100 fêmeas e quatro machos eram utilizados em cada geração, por exemplo, o aumento médio por geração na consanguinidade seria (4 + 100) ÷ (8)(4)(100) = 0,0325. Numa base anual, assumindo um intervalo de geração de cinco anos, isto equivaleria a um aumento médio anual da consanguinidade de 0,0065 ou 0,65 por cento.
Apesar dos resultados geralmente pobres obtidos com a consanguinidade, é uma ferramenta muito útil na criação de animais. A consanguinidade é essencial para o desenvolvimento de animais prepotentes – animais que uniformemente “carimbam” as suas características na sua progenitura. Porque a consanguinidade causa um aumento na proporção de genes semelhantes (bons ou maus, recessivos ou dominantes), as células reprodutivas do animal consanguíneo serão mais uniformes na sua composição genética. Quando esta uniformidade envolve um número relativamente grande de genes dominantes, a progenitura desse indivíduo exibirá uniformemente as características dominantes desse progenitor.
A consanguinidade também pode ser usada para descobrir genes que produzem anormalidades ou morte – genes que, nos rebanhos de consanguinidade, estão geralmente presentes em baixas frequências. Estes genes nocivos são quase sempre recessivos na sua natureza genética e os seus efeitos são ocultados ou mascarados pelos seus homólogos dominantes (alelos). Com excepção dos traços ligados ao sexo, os genes recessivos não são expressos se transportados isoladamente. Para que os seus efeitos se manifestem, devem estar presentes em duplicado. A probabilidade de estarem presentes em duplicado aumenta com a consanguinidade, porque a consanguinidade aumenta a proporção de genes semelhantes (tanto bons como maus) na população consanguínea. Com os efeitos destes genes descobertos, o criador pode eliminá-los do seu rebanho. Ele abateria a prole que mostrasse o efeito indesejável destes genes recessivos e também abateria os pais que são portadores dos genes indesejáveis. Além disso, dois terços da progenitura “normal” destes progenitores portadores são, eles próprios, portadores destes mesmos genes indesejáveis. Na ausência de testes de reprodução para separar os portadores dos não portadores, seria também necessário abater todos os progenitores “normais” dos progenitores portadores.
Os criadores podem utilizar um teste de consanguinidade para identificar portadores de genes autossómicos recessivos nocivos (como os responsáveis pelo ronco anão no gado, pela hiperostose nos suínos, ou pelo criptorcidismo nos ovinos). Um teste de consanguinidade verifica apenas os genes recessivos que o animal testado (geralmente o macho) transporta. O exemplo seguinte e os números dados são pertinentes para o gado bovino, onde uma prole por gestação é habitual. Se quisermos ter a certeza ao nível 0,01 de probabilidade de um touro estar livre de genes recessivos autossómicos nocivos, teríamos de o acasalar a pelo menos 35 das suas filhas. Ele é acasalado a mais de 35 das suas filhas porque apenas metade delas são esperadas para carregar qualquer gene recessivo nocivo que o seu reprodutor carregue. Precisamos da produção de 35 bezerros normais sem um único bezerro anormal para mostrar o touro livre (ao nível de probabilidade 0,01) de qualquer gene autossómico recessivo nocivo.
Testes em ovinos e suínos exigiriam acasalamentos a menos filhas. Os números reais dependeriam do número médio de descendência produzida por gestação em cada espécie. Utilizando o nível de probabilidade 0,01 e assumindo que o tamanho médio da ninhada é de 1,5 em ovelhas e 8,0 em suínos, o número de acasalamentos de reprodutoras necessárias seria cerca de 24+ para ovelhas e 5+ para suínos.
Outra utilização importante da consanguinidade está no desenvolvimento de famílias distintas ou linhas consanguíneas. Começando com uma população genética inicialmente diversificada, a consanguinidade resulta na formação de várias linhas, cada uma diferindo geneticamente da outra. A consanguinidade contínua dentro destas linhas tende a alterar a frequência de alguns dos genes encontrados na população inicial. Por exemplo, se um determinado gene estiver presente em apenas 1% dos animais da população inicial, a consanguinidade e o desenvolvimento de linhas distintas pode resultar na presença deste gene em todos ou quase todos os animais de algumas linhas e em nenhum ou apenas alguns dos animais de outras linhas. As linhagens consanguíneas são utilizadas de várias maneiras, mas são provavelmente mais utilizadas no desenvolvimento de frangos híbridos ou milho híbrido de semente.
Uma forma geralmente suave de consanguinidade (linebreeding) está a ser utilizada com sucesso por alguns produtores de sementes e produtores comerciais. O seu objectivo é manter um elevado grau de relação entre os animais do rebanho e alguns antepassados ou antepassados notáveis. Com a consanguinidade em geral, não há nenhuma tentativa de aumentar a relação entre a descendência e qualquer antepassado em particular. Num programa de consanguinidade há uma tentativa deliberada de manter ou aumentar a relação entre a descendência e um antepassado (ou antepassados) admirado específico. Esta característica distingue a consanguinidade como uma forma especial de consanguinidade.
O coeficiente de consanguinidade da descendência produzida num programa de consanguinidade é geralmente baixo, mas depende do tipo de programa de reprodução seguido. A figura 5 ilustra dois programas de reprodução cruzada. O programa de reprodução delineado na parte A mostra uma linha directa de relação entre a descendência “x” e o antepassado desejado “5”. Apenas um nível suave de consanguinidade no animal “x” (Fx = 0,03125) é alcançado. O coeficiente de relação obtido entre o animal “x” e o animal “5” é de 0,2462. A parte B é um programa de consanguinidade baseado em acasalamentos contínuos entre o animal “x” e “S” de 0,375 e um coeficiente de relação entre os animais “x” e “S” de 0,78.
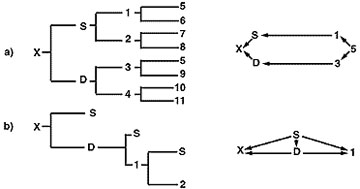 Figura 5
Figura 5
Diagramas de pedigree e seta para dois programas de reprodução em linha.
O programa de criação de reprodutores na parte B concentra efectivamente genes do animal “S”, mas devido ao rápido aumento da consanguinidade da progenitura produzida neste programa, o criador corre o risco de ter um desempenho muito reduzido e uma elevada probabilidade de defeitos genéticos. Os programas de consanguinidade são melhor utilizados em rebanhos de raça pura e de elevado desempenho quando um indivíduo verdadeiramente superior geneticamente nesse rebanho tiver sido identificado e avaliado por testes de descendência. A concentração dos genes desse indivíduo seria então melhor realizada através do acasalamento com fêmeas não relacionadas para reduzir o risco de efeitos nocivos associados a uma consanguinidade tão intensa.
Resumo
A consanguinidade é tecnicamente definida como o acasalamento de animais mais próximos do que a relação média dentro da raça ou população em questão. Para efeitos práticos, se dois indivíduos acasalados não tiverem antepassados comuns nas últimas cinco ou seis gerações, a sua progenitura seria considerada como sendo um surto. A consequência genética primária da consanguinidade é aumentar a frequência do acasalamento de genes semelhantes. Todas as alterações genéticas e fenotípicas associadas com a prática da consanguinidade derivam desta única consequência primária. Em geral, a consanguinidade resulta numa diminuição geral do desempenho. É mais obviamente reflectida numa menor eficiência reprodutiva, incluindo taxas de mortalidade mais elevadas, taxas de crescimento mais baixas e uma maior frequência de defeitos hereditários. Apesar destes efeitos geralmente nocivos, a consanguinidade é um instrumento muito útil no campo da reprodução animal. Permite ao criador descobrir e eliminar genes recessivos nocivos dentro da população. É também essencial para o desenvolvimento de animais prepotentes e é desejável no desenvolvimento de linhas familiares distintas. Além disso, os produtores de sementes e comerciais têm utilizado com sucesso a linebreza para manter um grau de relação genética nos seus animais com algum antepassado ou antepassados excepcionais.
Glossário
Terms descrevem vários esquemas de acasalamento e a descendência resultante de programas específicos de acasalamento.
- Backcross
Progenia resultante do acasalamento de um animal cruzado de dois cruzamentos com uma das raças parentais. Por exemplo, utilizando duas raças designadas como P1, e P2, a progênie backcross seria produzida pelo acasalamento do animal cruzado de duas raças (Pl × P2) com uma das raças parentais P1 ou P2. - Crisscrossing
Um programa contínuo de cruzamentos em que há um uso alternativo de machos pertencentes a duas raças. Utilizando duas raças designadas como P1 e P2, um programa de cruzamentos, começando com o cruzamento de duas raças um imal (P1 × P2), começaria por cruzamentos com uma das raças parentais. As fêmeas resultantes destes cruzamentos seriam criadas a um macho P2, × P2, e assim por diante. - Crossbred=br>Progenia resultante do acasalamento de animais outcross pertencentes a diferentes raças.
- Classificação
Acoplamento de machos de raça pura de uma dada raça a fêmeas não de raça pura e a resultante progenitura feminina em gerações sucessivas. - Linha de animais produzidos por animais relacionados com o acasalamento.
- Consanguinidade
Acoplamento de animais mais próximos uns dos outros do que a relação média dentro da raça ou população em causa. - Incrossbred
Progenia resultante do acasalamento de animais de linhagens consanguíneas de diferentes raças. - Linebreza
Forma geralmente suave de consanguinidade em que os animais acasalados estão relacionados com algum indivíduo supostamente proeminente. - Topcross
Progenia resultante do acasalamento de machos consanguíneos com fêmeas não consanguíneas de outra raça. - Topincross
Progenia resultante do acasalamento de machos consanguíneos com fêmeas não consanguíneas da mesma raça. - Cruzamento de duas raças
Progenia resultante do acasalamento de machos de uma raça com fêmeas de outra raça.
li>cruzamento de animais pertencentes a diferentes géneros (por exemplo, acasalamento de gado doméstico, Bos taurus ou Bos indicus, ao búfalo americano, Bisontes).
Incross>br>Progenia resultante do acasalamento de animais de diferentes linhagens consanguíneas dentro de uma raça.
Sanguinidade
Acasalamento de animais menos relacionados entre si do que a relação média dentro da raça ou população em questão.Encastramento
Progenia resultante do acasalamento de animais não relacionados dentro de uma raça. Cruzamento de espécies
Acoplamento de animais pertencentes a espécies diferentes (por exemplo acasalamento de gado de raça europeia, Bos taurus, a gado Brahman, Bos indicus). Cruzamento de três raças em rotação
Um programa contínuo de cruzamentos em que machos de três raças são utilizados em rotação. Utilizando três raças designadas como P1, P2 e P3, a primeira geração envolveria a produção de cruzamentos de dois machos, P1 x P2. Na segunda geração, as fêmeas de cruzamentos de duas raças seriam cruzadas com machos da terceira raça, (P1 × P2) × P3; as fêmeas de cruzamentos de três raças seriam cruzadas com machos de uma das raças utilizadas para produzir os cruzamentos de duas raças, × P1, e assim por diante.li> Topcross
Progenia resultante do acasalamento de animais pertencentes a diferentes famílias dentro de uma raça.