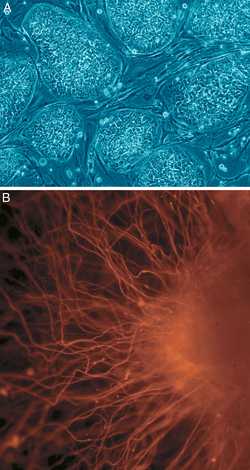
B: Células nerviosas
En biología celular, pluripotencia (Lat. pluripotentia, «capacidad para muchos») se refiere a una célula madre que tiene el potencial de diferenciarse en cualquiera de las tres capas germinales: endodermo (revestimiento interior del estómago, tracto gastrointestinal, los pulmones), mesodermo (músculo, hueso, sangre, urogenital), o ectodermo (tejidos epidérmicos y sistema nervioso), pero no en tejidos extraembrionarios como la placenta. Sin embargo, la pluripotencia celular es un continuo, que va desde la célula completamente pluripotente que puede formar todas las células del embrión propiamente dicho, por ejemplo, las células madre embrionarias y las iPSC (véase más adelante), hasta la célula incompleta o parcialmente pluripotente que puede formar células de las tres capas germinales pero que puede no presentar todas las características de las células completamente pluripotentes.
Pluripotencia inducidaEditar
Las células madre pluripotentes inducidas, comúnmente abreviadas como células iPS o iPSCs, son un tipo de célula madre pluripotente derivada artificialmente de una célula no pluripotente, típicamente una célula somática adulta, mediante la inducción de una expresión «forzada» de ciertos genes y factores de transcripción. Estos factores de transcripción desempeñan un papel clave en la determinación del estado de estas células y también pone de manifiesto el hecho de que estas células somáticas sí conservan la misma información genética que las células embrionarias tempranas. La capacidad de inducir células a un estado pluripotente fue inicialmente pionera en 2006 utilizando fibroblastos de ratón y cuatro factores de transcripción, Oct4, Sox2, Klf4 y c-Myc; esta técnica, denominada reprogramación, les valió posteriormente a Shinya Yamanaka y John Gurdon el Premio Nobel de Fisiología o Medicina. A continuación, en 2007, se indujeron con éxito iPSC humanas derivadas de fibroblastos dérmicos humanos utilizando métodos similares a los utilizados para la inducción de células de ratón. Estas células inducidas presentan rasgos similares a los de las células madre embrionarias (CME), pero no requieren el uso de embriones. Algunas de las similitudes entre las ESC y las iPSC incluyen la pluripotencia, la morfología, la capacidad de autorrenovación, un rasgo que implica que pueden dividirse y replicarse indefinidamente, y la expresión génica.
También se cree que los factores epigenéticos están implicados en la reprogramación real de las células somáticas para inducir la pluripotencia. Se ha teorizado que ciertos factores epigenéticos podrían funcionar para eliminar las marcas epigenéticas somáticas originales con el fin de adquirir las nuevas marcas epigenéticas que forman parte del logro de un estado pluripotente. La cromatina también se reorganiza en las iPSC y se asemeja a la que se encuentra en las CME en el sentido de que está menos condensada y, por tanto, es más accesible. Las modificaciones de la eucromatina también son comunes, lo que también es coherente con el estado de la eucromatina que se encuentra en las CME.
Debido a su gran similitud con las CME, las iPSC han sido de gran interés para la comunidad médica y de investigación. Las iPSC podrían tener potencialmente las mismas implicaciones y aplicaciones terapéuticas que las CME pero sin el controvertido uso de embriones en el proceso, un tema de gran debate bioético. De hecho, la pluripotencia inducida de células somáticas en células iPS indiferenciadas fue aclamada en un principio como el fin del controvertido uso de células madre embrionarias. Sin embargo, se descubrió que las iPSC eran potencialmente tumorigénicas y, a pesar de los avances, nunca fueron aprobadas para la investigación en fase clínica en Estados Unidos. También se han encontrado contratiempos como las bajas tasas de replicación y la senescencia temprana al fabricar iPSCs, lo que dificulta su uso como sustitutas de las ESCs.
Además, se ha determinado que la expresión somática de factores de transcripción combinados puede inducir directamente otros destinos celulares somáticos definidos (transdiferenciación); los investigadores identificaron tres factores de transcripción específicos del linaje neural que podían convertir directamente los fibroblastos de ratón (células de la piel) en neuronas totalmente funcionales. Este resultado desafía la naturaleza terminal de la diferenciación celular y la integridad del compromiso de linaje; e implica que, con las herramientas adecuadas, todas las células son totipotentes y pueden formar todo tipo de tejidos.
Algunos de los posibles usos médicos y terapéuticos de las iPSCs derivadas de pacientes incluyen su uso en trasplantes de células y tejidos sin el riesgo de rechazo que se encuentra habitualmente. Las iPSC pueden sustituir potencialmente a los modelos animales inadecuados, así como a los modelos in vitro utilizados para la investigación de enfermedades.

Estados de pluripotencia ingenua vs. primadaEditar
Los hallazgos recientes con respecto a los epiblasto antes y después de la implantación han producido propuestas para clasificar la pluripotencia en dos fases distintas: «naive» y «primed». Las células madre primitivas que se utilizan habitualmente en la ciencia y que se denominan células madre embrionarias (ESC) se derivan de un epiblasto previo a la implantación; dicho epiblasto es capaz de generar todo el feto, y una célula del epiblasto es capaz de contribuir a todos los linajes celulares si se inyecta en otro blastocisto. Por otra parte, se observan varias diferencias marcadas entre los epiblasto pre y post implantación, como su diferencia en la morfología, en la que el epiblasto después de la implantación cambia su morfología a una forma parecida a una copa llamada «cilindro de huevo», así como la alteración cromosómica en la que uno de los cromosomas X sufre una inactivación aleatoria en la fase temprana del cilindro de huevo, conocida como inactivación del X. Durante este desarrollo, las células del epiblasto del cilindro del huevo son sistemáticamente dirigidas por factores de crecimiento de fibroblastos, señalización Wnt y otros factores inductivos a través del saco vitelino circundante y el tejido del trofoblasto, de tal manera que se vuelven instructivamente específicas según la organización espacial.
Otra diferencia importante que se observó, con respecto a la potencia celular, es que las células madre del epiblasto post-implantación son incapaces de contribuir a las quimeras del blastocisto, lo que las distingue de otras células madre pluripotentes conocidas. Las líneas celulares derivadas de dichos epiblasto post-implantación se denominan células madre derivadas del epiblasto, que se obtuvieron por primera vez en el laboratorio en 2007; a pesar de su nomenclatura, que tanto las ESC como las EpiSC se derivan de epiblasto, sólo que en fases diferentes del desarrollo, y que la pluripotencia sigue intacta en el epiblasto post-implantación, como demuestra la expresión conservada de Nanog, Fut4 y Oct-4 en las EpiSC, hasta la somitogénesis y puede revertirse a mitad de camino mediante la expresión inducida de Oct-4.