Dale Vogt, Helen A. Swartz y John Massey
Departamento de Ciencias Animales
Los diversos esquemas de apareamiento de los animales se clasifican en dos grandes categorías – endogamia y exogamia. La clasificación depende de la cercanía de la relación biológica entre los compañeros. Dentro de cada categoría, existe una amplia variación en la intensidad de esta relación. Una línea muy fina separa las dos categorías. El apareamiento de animales estrechamente emparentados (por ejemplo, padre e hijo, hermano y hermana o medio hermano) es endogámico. Con animales menos emparentados (primos hermanos, primos segundos), la gente no está de acuerdo sobre dónde trazar la línea entre la endogamia y la exogamia.
Técnicamente, la endogamia se define como el apareamiento de animales más emparentados que la relación media dentro de la raza o población en cuestión. Los apareamientos entre animales menos emparentados que esto, por tanto, constituirían una superación. En el cuadro 1 se describen estos dos sistemas de apareamiento, con distintas intensidades en cada uno de ellos. Los apareamientos indicados dentro de la categoría de endogamia se explican por sí mismos; los de la categoría de exogamia se definen en el glosario.
Tabla 1
Grados de endogamia y exogamia ordenados según la relación biológica entre las parejas indicadas. (Al leer de arriba a abajo, la relación biológica entre los compañeros disminuye constantemente.)
| Entrecruzamiento o exogamia | Coeficiente de relación entre compañeros | Relación biológica entre compañeros |
|---|---|---|
| Entrecruzamiento | 50 por ciento | Padre × descendiente; hermanos completos |
| Semejanza | 25 por ciento | Medio hermanos; primos hermanos dobles; tía × sobrino; tío × sobrina |
| Entrecruzamiento | 12-1/2 por ciento | Primos hermanos |
| Encruzamiento | 6-1/4 por ciento | Primos segundos |
| Encruzamiento | ? | Cruzamiento en línea1 |
| Cruzamiento interno/externo | 0 por ciento | Cruzamiento aleatorio dentro de la raza o población2 |
| Superación | 0 por ciento | Cruzamiento |
| Superación | 0 por ciento | Cruzamiento de razas | Cruzamiento de razas | 0 por ciento | Cruzamiento de especies |
| Cruzamiento de razas | 0 por ciento | Cruzamiento de géneros |
1En un programa de cría en línea, el coeficiente de relación entre compañeros suele ser bajo; sin embargo, puede ser muy variable.
2El apareamiento aleatorio dentro de una raza o población significa que las parejas se eligen por azar. Debe entenderse que bajo esta circunstancia es posible que se produzca endogamia o exogamia.
Relaciones biológicas entre animales
Se considera que los individuos están biológicamente relacionados cuando tienen uno o más ancestros comunes. A efectos prácticos, si dos individuos no tienen ningún antepasado común en las últimas cinco o seis generaciones, se considera que no están emparentados.
El parentesco biológico es importante en la cría de animales porque cuanto más cercano sea el parentesco, mayor será el porcentaje de genes afines que lleven los dos individuos. La cercanía del parentesco está determinada por tres factores:
- Cuánto tiempo atrás en los pedigríes de los dos animales aparece el ancestro común
- Cuántos ancestros comunes tienen
- Cuán frecuentemente aparecen los ancestros comunes. También influye la posible consanguinidad del ancestro o ancestros comunes
Medida del grado de parentesco biológico
El coeficiente de parentesco es un único valor numérico que considera todos los factores mencionados. Es una medida del grado de similitud de los genotipos (constituciones genéticas) de los dos animales. Se estima mediante la expresión:
RBC = sigma ÷ Raíz cuadrada de (1 + FB)(1 + FC)
Ecuación 1 donde:
RBC = el coeficiente de relación entre los animales B y C que queremos medir.
sigma = el símbolo griego que significa «suma».
(1/2) = la fracción del material genético de un individuo que se transmite a su progenie. Se utiliza en el cálculo del coeficiente de parentesco porque representa la probabilidad de que, en una generación cualquiera, un gen idéntico de un par de genes dado se transmita a cada una de dos progenies concretas. También es la probabilidad de que un gen distinto de un par de genes dado se transmita a las dos progenies.
n = el número de gneraciones entre el animal B y el ancestro común.
n’ = el número de generaciones entre el animal C y el ancestro común
FA, FB, FC = coeficientes de consanguinidad del ancestro común y de los animales B y C, respectivamente.
Si ninguno de los animales es consanguíneo, el coeficiente de parentesco se estima como:
RBC = sigma
Ecuación 2
El uso de esta expresión se puede demostrar con el pedigrí de muestra de hermanos completos y el diagrama de flechas (Figura 1). En este ejemplo, se supone que ni el padre ni la madre son consanguíneos. El diagrama de flechas de la derecha muestra las trayectorias del flujo de genes desde cada uno de los ancestros comunes (D y E) hasta los animales cuyo coeficiente de parentesco estamos midiendo (B y C).
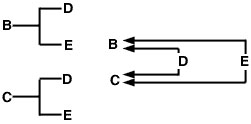
Figura 1
El problema ahora es trazar todas las trayectorias posibles desde el animal B hasta el animal C que pasan por un ancestro común. En este caso, hay dos de estos caminos (Figura 2).
Figura 2
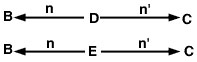
Ecución 3
Como hemos supuesto que no hay consanguinidad en este ejemplo, el coeficiente de relación entre los animales B y C se estima como:
RBC = sigma = (1/2)1 + 1 + (1/2)1 + 1 = (1/2)2 + (1/2)2 = 0.50
Utilidad de la información del coeficiente de relación
Un ganadero encontraría valiosa la información del coeficiente de relación en varias situaciones. Puede, por ejemplo, querer vender un animal emparentado con otro que previamente se ha vendido a un precio elevado. Cuanto mayor sea el coeficiente de relación entre ambos, mejor será su uso como punto de venta. O bien, puede querer comprar uno de dos toros emparentados y uno de ellos puede costar más de lo que desea pagar. Si el coeficiente de relación entre los dos toros es alto, podría estar tan bien con el toro de menor precio como con el más caro.
Un uso práctico del coeficiente de relación es estimar el valor de rendimiento de un animal no probado. Para estimar el valor, debemos conocer el valor de rendimiento de un animal relacionado, el coeficiente de relación entre los animales probados y no probados, y el valor de rendimiento medio de la raza, el rebaño o el grupo al que pertenecen los animales probados y no probados.
Como ejemplo, considere un rebaño con una ganancia media diaria (ADG) en el cebadero de 2,25 libras por día. Supongamos además que un padre de este rebaño tuvo una ADG de 3,50 libras por día en el corral de engorde, mientras que un medio hermano más joven no ha sido evaluado aún para la ADG en el corral de engorde. Asumiendo que no hay consanguinidad, el coeficiente de relación entre los medios hermanos es de 0,25. La mejor estimación del ADG en corral del animal no evaluado es que se desviará de la media del rebaño en un 25 por ciento como lo hace el valor de rendimiento del medio hermano evaluado. Utilizando estas cifras, el valor más probable de ADG en el cebadero del animal no probado es 2,25 + (0,25) × (3,50 – 2,25) o 2,56 libras por día.
Medida del grado de consanguinidad
Cuando calculamos un coeficiente de consanguinidad, estamos intentando medir la reducción porcentual probable en la frecuencia de emparejamiento de genes disímiles (reducción de la heterocigosidad). Esta reducción es relativa a una población base. La población base suele ser la raza en cuestión en una fecha a la que se remontan los pedigríes. Se supone que los animales de esta población base no son consanguíneos. Esto no significa que estos animales de la población base tengan genes distintos en cada par. No hay forma de saber cuántos de sus pares de genes consistían en genes similares o disímiles. El coeficiente de consanguinidad que se calcula es simplemente relativo a esa base y refleja la probable reducción porcentual de cuantos pares de genes disímiles tenían los animales promedio de la población base.
La expresión general para determinar el coeficiente de consanguinidad es:
FX = sigma
Ecuación 4 donde:
FX = el coeficiente de consanguinidad del animal X.
sigma = el símbolo griego que significa «suma».
(1/2) = la fracción del material genético de un individuo que se transmite a su progenie. Se utiliza en el cálculo del coeficiente de parentesco porque representa la probabilidad de que, en una generación cualquiera, un gen idéntico de un par de genes dado se transmita a cada una de dos progenies concretas. También es la probabilidad de que un gen distinto de un par de genes dado se transmita a las dos progenies.
n = el número de generaciones entre el animal B y el ancestro común.
n’ = el número de generaciones entre el animal C y el ancestro común.
+1 = se añade a n y n’ para tener en cuenta la generación adicional entre el animal × y sus padres.
FA = el coeficiente de consanguinidad del ancestro común.
Si ninguno de los padres es consanguíneo, pero si están emparentados, el coeficiente de consanguinidad de su progenie es la mitad de su coeficiente de parentesco: 1/2 RBC Esto se puede demostrar utilizando un apareamiento de hermanos completos para facilitar la comparación con el coeficiente de parentesco de los hermanos completos calculado anteriormente. En este caso, el pedigrí del animal X y el diagrama de flechas serán los siguientes:
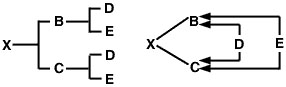
Figura 3
El problema ahora es trazar todos los caminos posibles desde el padre (B) a la madre (C) a través de cada ancestro común. Al igual que en el problema del coeficiente de parentesco, hay dos de estos caminos:
![]()
Figura 4
Como hemos asumido que ninguno de los padres es consanguíneo, el coeficiente de consanguinidad del animal × se estima como:
FX = sigma = (1/2)1 + 1 + 1 + (1/2)1 + 1 = (1/2)3 + (1/2)3 = 0.25
Consecuencia 5
Esto es la mitad del coeficiente de relación entre hermanos completos cuando no hay consanguinidad.
Consecuencias genéticas de la consanguinidad
La consecuencia genética básica de la consanguinidad es promover lo que se conoce técnicamente como homocigosidad. Esto significa que hay un aumento en la frecuencia de emparejamiento de genes similares. Acompañando a este aumento, debe haber una disminución en la frecuencia de emparejamiento de genes disímiles. Esto se denomina disminución de la heterocigosidad. Estos eventos simultáneos son las razones subyacentes de los efectos generales sobre el rendimiento que observamos con la endogamia.
Razones para la endogamia
Es posible desarrollar líneas endogámicas altamente productivas de ganado doméstico. Sin embargo, hasta la fecha, estos intentos han tenido poco éxito aparente. Aunque ocasionalmente se producen animales de alto rendimiento, la endogamia suele provocar una reducción general del rendimiento. Esta reducción se manifiesta de muchas maneras. Los efectos más obvios de la consanguinidad son una menor eficiencia reproductiva, incluyendo mayores tasas de mortalidad, menores tasas de crecimiento y una mayor frecuencia de anormalidades hereditarias. Así lo han demostrado numerosos estudios con ganado vacuno, caballos, ovejas, cerdos y animales de laboratorio.
La magnitud de esta disminución del rendimiento, en general, es proporcional al grado de consanguinidad. Cuanto mayor sea el grado de consanguinidad, mayor será la reducción del rendimiento. La reducción real del rendimiento no es la misma en todas las especies ni en todos los rasgos. Algunas características (como la calidad de la carne) apenas se ven influidas por la consanguinidad; otras (como la eficiencia reproductiva) se ven muy influidas por la consanguinidad. Por lo tanto, no podemos hacer una afirmación generalizada sobre la cantidad de reducción en el «rendimiento» que resultaría de una cantidad específica de consanguinidad y esperar que sea aplicable en una amplia variedad de situaciones.
Sin embargo, es posible predecir la magnitud del efecto de la consanguinidad en rasgos específicos. Dichas predicciones se basan en los resultados obtenidos realmente en condiciones experimentales en las que se han alcanzado diversos niveles de consanguinidad. En una investigación con cerdos llevada a cabo en el Midwest Regional Swine Breeding Laboratory, Dickerson y otros (1954) señalan que por cada aumento del 10% en la consanguinidad (de los cerdos de la camada), se produce una disminución de 0,20, 0,35, 0,38 y 0,44 cerdos por camada al nacimiento, a los 21 días, a los 56 días y a los 154 días, respectivamente. Podemos utilizar estas cifras para proporcionar estimaciones de las disminuciones esperadas en el tamaño de la camada (a edades comparables de la camada) en otras piaras de cerdos.
La mayoría de los estudios de consanguinidad sugieren que cada incremento sucesivo de la unidad de consanguinidad resulta en una disminución proporcional del rendimiento. Las estimaciones de los incrementos medios del porcentaje de consanguinidad dentro de una piara cerrada pueden realizarse con la expresión:
Incremento medio de consanguinidad aumento de consanguinidad = (Número de machos + Número de hembras) ÷ (8)(Número de machos)(Número de hembras)
En un rebaño cerrado de ganado en el que se utilizaran 100 hembras y cuatro machos en cada generación, por ejemplo, el aumento medio por generación de la consanguinidad sería (4 + 100) ÷ (8)(4)(100) = 0,0325. Sobre una base anual, suponiendo un intervalo generacional de cinco años, esto equivaldría a un aumento medio anual de la consanguinidad de 0,0065 o 0,65 por ciento.
A pesar de los resultados generalmente pobres obtenidos con la consanguinidad, es una herramienta muy útil en la cría de animales. La consanguinidad es esencial para el desarrollo de animales prepotentes, es decir, animales que «imprimen» de manera uniforme sus características a su progenie. Como la consanguinidad provoca un aumento de la proporción de genes afines (buenos o malos, recesivos o dominantes), las células reproductoras del animal consanguíneo serán más uniformes en su composición genética. Cuando esta uniformidad implica un número relativamente grande de genes dominantes, la progenie de ese individuo mostrará de manera uniforme las características dominantes de ese progenitor.
La endogamia también puede utilizarse para descubrir genes que producen anormalidades o la muerte, genes que, en los rebaños de cría, suelen estar presentes en bajas frecuencias. Estos genes dañinos son casi siempre de naturaleza genética recesiva y sus efectos quedan ocultos o enmascarados por sus homólogos dominantes (alelos). Salvo en el caso de los rasgos ligados al sexo, los genes recesivos no se expresan si se portan solos. Para que sus efectos se manifiesten, deben estar presentes por duplicado. La probabilidad de que estén presentes por duplicado aumenta con la endogamia, ya que ésta aumenta la proporción de genes similares (tanto buenos como malos) en la población endogámica. Una vez descubiertos los efectos de estos genes, el criador puede eliminarlos de su rebaño. Eliminaría la progenie que mostrara el efecto indeseable de estos genes recesivos y también eliminaría a los padres portadores de los genes indeseables. Además, se espera que dos tercios de la descendencia «normal» de estos padres portadores sean a su vez portadores de estos mismos genes indeseables. En ausencia de pruebas de cría para separar a los portadores de los no portadores, también sería necesario sacrificar a toda la progenie «normal» de los padres portadores.
Los criadores pueden utilizar una prueba de consanguinidad para identificar a los portadores de genes autosómicos recesivos perjudiciales (como los responsables del enanismo del roncador en el ganado vacuno, la hiperostosis en el ganado porcino o la criptorquidia en el ganado ovino). Una prueba de endogamia comprueba sólo los genes recesivos de los que es portador el animal analizado (normalmente el macho). El siguiente ejemplo y los números que se dan son pertinentes para el ganado vacuno, donde lo habitual es una cría por gestación. Si queremos estar seguros, con un nivel de probabilidad del 0,01, de que un toro está libre de genes recesivos autosómicos dañinos, tendríamos que aparearlo con al menos 35 de sus hijas. Se aparea con más de 35 de sus hijas porque se espera que sólo la mitad de ellas sean portadoras de cualquier gen recesivo dañino que lleve su padre. Necesitamos la producción de 35 terneros normales sin un solo ternero anormal para demostrar que el toro está libre (a un nivel de probabilidad de 0,01) de cualquier gen autosómico recesivo dañino.
Las pruebas en ovejas y cerdos requerirían apareamientos con menos hijas. Las cifras reales dependerían del número medio de progenie producida por gestación en cada especie. Utilizando el nivel de probabilidad de 0,01 y asumiendo que el tamaño medio de la camada es de 1,5 en ovejas y de 8,0 en cerdos, el número de apareamientos padre-hija necesarios sería de unos 24+ en ovejas y 5+ en cerdos.
Otro uso importante de la consanguinidad es el desarrollo de familias distintas o líneas consanguíneas. Partiendo de una población genética inicialmente diversa, la consanguinidad da lugar a la formación de varias líneas, cada una de las cuales difiere genéticamente de la otra. La endogamia continuada dentro de estas líneas tiende a cambiar la frecuencia de algunos de los genes encontrados en la población inicial. Por ejemplo, si un gen concreto está presente sólo en el 1% de los animales de la población inicial, la endogamia y el desarrollo de líneas distintas pueden hacer que este gen esté presente en todos o casi todos los animales de algunas líneas y en ninguno o sólo en unos pocos de los animales de otras líneas. Las líneas consanguíneas se utilizan de varias maneras, pero probablemente sean las más utilizadas en el desarrollo de pollos híbridos o de semillas de maíz híbridas.
Una forma generalmente suave de endogamia (linebreeding) está siendo utilizada con éxito por algunos productores de semillas y comerciales. Su objetivo es mantener un alto grado de relación entre los animales del rebaño y algún ancestro o ancestros destacados. Con la consanguinidad en general, no se intenta aumentar el parentesco entre la descendencia y algún ancestro en particular. En un programa de cría en línea hay un intento deliberado de mantener o aumentar la relación entre la descendencia y un ancestro (o ancestros) específico admirado. Esta característica distingue a la cría en línea como una forma especial de endogamia.
El coeficiente de endogamia de la descendencia producida en un programa de cría en línea suele ser bajo, pero depende del tipo de programa de cría que se siga. La figura 5 ilustra dos programas de cría en línea. El programa de cría descrito en la parte A muestra una línea de parentesco directa entre la descendencia «x» y el ancestro deseado «5». Sólo se alcanza un nivel leve de consanguinidad en el animal «x» (Fx = 0,03125). El coeficiente de parentesco obtenido entre el animal «x» y el animal «5» es de 0,2462. La parte B es un programa de cría en línea basado en apareamientos continuos padre-hija que, tras sólo dos generaciones, produce un nivel de consanguinidad en el animal «x» de 0,375 y un coeficiente de relación entre los animales «x» y «S» de 0,78.
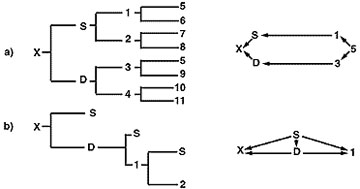 Figura 5
Figura 5
Pedigrees y diagramas de flechas para dos programas de cría en línea.
El programa padre-hija de la parte B concentra eficazmente los genes del animal «S», pero debido al rápido aumento de la consanguinidad de la progenie producida en este programa, el criador corre el riesgo de un rendimiento muy reducido y una alta probabilidad de defectos genéticos. Los programas de cría en línea se utilizan mejor en los rebaños de raza pura y de alto rendimiento cuando se ha identificado y evaluado mediante pruebas de progenie un individuo verdaderamente superior desde el punto de vista genético en ese rebaño. La mejor manera de concentrar los genes de ese individuo es apareándolo con hembras no emparentadas para reducir el riesgo de efectos nocivos asociados a una endogamia tan intensa.
Resumen
La endogamia se define técnicamente como el apareamiento de animales más emparentados que la relación media dentro de la raza o población en cuestión. A efectos prácticos, si dos individuos apareados no tienen un ancestro común en las últimas cinco o seis generaciones, su progenie se considerará ajena a la raza. La principal consecuencia genética de la consanguinidad es el aumento de la frecuencia de emparejamiento de genes similares. Todos los cambios genéticos y fenotípicos asociados a la práctica de la endogamia se derivan de esta consecuencia primaria. En general, la consanguinidad provoca una disminución global del rendimiento. Se refleja de forma más evidente en una menor eficiencia reproductiva, incluyendo mayores tasas de mortalidad, menores tasas de crecimiento y una mayor frecuencia de defectos hereditarios. A pesar de estos efectos generalmente perjudiciales, la consanguinidad es una herramienta muy útil en el campo de la cría de animales. Permite al criador descubrir y eliminar genes recesivos perjudiciales dentro de la población. También es esencial para el desarrollo de animales prepotentes y es deseable en el desarrollo de líneas familiares distintas. Además, los productores de semillas y comerciales han utilizado con éxito la cría en línea para mantener un grado de relación genética en sus animales con algún ancestro o ancestros destacados.
Glosario
Los términos describen varios esquemas de apareamiento y la progenie resultante de programas de apareamiento específicos.
- Cruzamiento a la inversa
Progenie resultante del apareamiento de un animal de cruce de dos razas con una de las razas parentales. Por ejemplo, utilizando dos razas designadas como P1, y P2, la progenie de retrocruzamiento se produciría apareando el animal de cruce de dos razas (Pl × P2) con cualquiera de las razas parentales P1 o P2. - Cruzamiento
Un programa continuo de cruces en el que hay un uso alternativo de machos pertenecientes a dos razas. Utilizando dos razas designadas como P1 y P2, un programa de entrecruzamiento, comenzando con el cruce de dos razas an imales (P1 × P2), comenzaría por el retrocruzamiento con una de las razas parentales . Las hembras resultantes de estos cruces se cruzarían con un macho P2, × P2, y así sucesivamente. - Cruzamiento
Progenie resultante del apareamiento de animales outcross pertenecientes a diferentes razas. - Cruzamiento de género
Cruzamiento de animales pertenecientes a diferentes géneros (por ejemplo, apareamiento de ganado doméstico, Bos taurus o Bos indicus, con el búfalo americano, Bison). - Clasificación
Clasificación de machos de raza pura de una determinada raza con hembras no puras y la descendencia femenina resultante en generaciones sucesivas. - Línea endogámica
Línea de animales producida por el apareamiento de animales emparentados. - Entrecruzamiento
Casamiento de animales más emparentados entre sí que el parentesco medio dentro de la raza o población de que se trate. - Cruzamiento
Procedencia resultante del apareamiento de animales de diferentes líneas endogámicas dentro de una raza. - Incruzamiento
Progenia resultante del apareamiento de animales de líneas consanguíneas de diferentes razas. - Linebreeding
Forma generalmente leve de endogamia en la que los animales apareados están emparentados con algún individuo supuestamente destacado. - Extractivismo
Aparejamiento de animales menos emparentados entre sí que la relación media dentro de la raza o población en cuestión. - Cruzamiento externo
Progenia resultante del apareamiento de animales no emparentados dentro de una raza. - Cruzamiento de especies
Cruzamiento de animales pertenecientes a especies diferentes (por ejemplo, apareamiento de ganado de raza europea, Bos taurus, con ganado Brahman, Bos indicus). - Cruzamiento rotativo de tres razas
Programa continuo de cruces en el que se utilizan machos de tres razas de forma rotativa. Utilizando tres razas designadas como P1, P2 y P3, la primera generación implicaría la producción de animales cruzados de dos razas, P1 x P2. En la segunda generación, las hembras cruzadas de dos razas se aparearían con machos de la tercera raza, (P1 × P2) × P3; las hembras cruzadas de tres razas se aparearían con machos de una de las razas utilizadas para producir los animales cruzados de dos razas, × P1, y así sucesivamente. - Topcross
Progenia resultante del apareamiento de animales pertenecientes a diferentes familias dentro de una raza. - Topcrossbred
Progenia resultante del apareamiento de machos consanguíneos con hembras no consanguíneas de otra raza. - Topincross
Progenia resultante del apareamiento de machos consanguíneos con hembras no consanguíneas de la misma raza. - Cruzamiento de dos razas
Progenia resultante del apareamiento de machos de una raza con hembras de otra raza.