Temperatura centralEditar

Los mamíferos regulan su temperatura central utilizando la información de los termorreceptores del hipotálamo, el cerebro, la médula espinal, los órganos internos y las grandes venas. Aparte de la regulación interna de la temperatura, puede entrar en juego un proceso llamado alostasis que ajusta el comportamiento para adaptarse al desafío de temperaturas extremas muy calientes o frías (y a otros desafíos). Estos ajustes pueden incluir la búsqueda de la sombra y la reducción de la actividad, o la búsqueda de condiciones más cálidas y el aumento de la actividad, o el acurrucamiento.La termorregulación del comportamiento tiene prioridad sobre la termorregulación fisiológica, ya que los cambios necesarios pueden verse afectados más rápidamente y la termorregulación fisiológica es limitada en su capacidad de respuesta a las temperaturas extremas.
Cuando la temperatura central cae, el suministro de sangre a la piel se reduce por una intensa vasoconstricción. El flujo sanguíneo hacia las extremidades (que tienen una gran superficie) se reduce de forma similar, y vuelve al tronco a través de las venas profundas que se encuentran junto a las arterias (formando venae comitantes). Esto actúa como un sistema de intercambio a contracorriente que cortocircuita el calor de la sangre arterial directamente en la sangre venosa que regresa al tronco, lo que provoca una pérdida mínima de calor en las extremidades cuando hace frío. Las venas subcutáneas de las extremidades están fuertemente constreñidas, lo que no sólo reduce la pérdida de calor de esta fuente, sino que también obliga a la sangre venosa a entrar en el sistema de contracorriente en las profundidades de las extremidades.
La tasa metabólica se incrementa, inicialmente por termogénesis sin escalofríos, seguida de termogénesis con escalofríos si las reacciones anteriores son insuficientes para corregir la hipotermia.
Cuando los termorreceptores detectan el aumento de la temperatura central, las glándulas sudoríparas de la piel se estimulan a través de los nervios simpáticos colinérgicos para secretar sudor sobre la piel, que, al evaporarse, enfría la piel y la sangre que fluye por ella. El jadeo es un efector alternativo en muchos vertebrados, que enfría el cuerpo también por la evaporación del agua, pero esta vez de las membranas mucosas de la garganta y la boca.
Glucosa en sangreEditar

Los niveles de azúcar en la sangre se regulan dentro de límites bastante estrechos. En los mamíferos, los principales sensores para ello son las células beta de los islotes pancreáticos. Las células beta responden a un aumento del nivel de azúcar en la sangre segregando insulina en la sangre y, al mismo tiempo, impidiendo que sus células alfa vecinas segreguen glucagón en la sangre. Esta combinación (niveles altos de insulina en sangre y niveles bajos de glucagón) actúa sobre los tejidos efectores, entre los que destacan el hígado, las células grasas y las células musculares. El hígado se inhibe de producir glucosa, tomándola en su lugar, y convirtiéndola en glucógeno y triglicéridos. El glucógeno se almacena en el hígado, pero los triglicéridos se segregan en la sangre en forma de partículas de lipoproteínas de muy baja densidad (VLDL) que son absorbidas por el tejido adiposo, donde se almacenan como grasas. Las células grasas captan la glucosa a través de transportadores especiales de glucosa (GLUT4), cuyo número en la pared celular aumenta como efecto directo de la acción de la insulina sobre estas células. La glucosa que entra en las células grasas de esta manera se convierte en triglicéridos (a través de las mismas vías metabólicas que utiliza el hígado) y luego se almacena en esas células grasas junto con los triglicéridos derivados de las VLDL que se produjeron en el hígado. Las células musculares también captan la glucosa a través de los canales de glucosa GLUT4, sensibles a la insulina, y la convierten en glucógeno muscular.
Una caída de la glucosa en sangre, hace que se detenga la secreción de insulina, y que se segregue glucagón desde las células alfa a la sangre. Esto inhibe la captación de glucosa de la sangre por el hígado, las células grasas y los músculos. En su lugar, el hígado se ve fuertemente estimulado para fabricar glucosa a partir del glucógeno (a través de la glucogenólisis) y de fuentes que no son carbohidratos (como el lactato y los aminoácidos desaminados) mediante un proceso conocido como gluconeogénesis. La glucosa así producida se descarga en la sangre corrigiendo el error detectado (hipoglucemia). El glucógeno almacenado en los músculos permanece en ellos y sólo se descompone, durante el ejercicio, en glucosa-6-fosfato y, de ahí, en piruvato para ser alimentado en el ciclo del ácido cítrico o convertido en lactato. Sólo el lactato y los productos de desecho del ciclo del ácido cítrico se devuelven a la sangre. El hígado puede tomar sólo el lactato y, mediante el proceso de gluconeogénesis que consume energía, convertirlo de nuevo en glucosa.
Niveles de hierroEditar
Regulación del cobre
Niveles de gases en sangreEditar
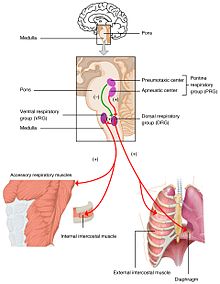
Los cambios en los niveles de oxígeno, dióxido de carbono y pH plasmático se envían al centro respiratorio, en el tronco cerebral, donde se regulan.La presión parcial de oxígeno y de dióxido de carbono en la sangre arterial es controlada por los quimiorreceptores periféricos (PNS) en la arteria carótida y el arco aórtico. Los quimiorreceptores centrales (SNC) de la médula oblonga del tronco cerebral detectan los cambios en la presión parcial del dióxido de carbono como una alteración del pH del líquido cefalorraquídeo. La información de estos conjuntos de sensores se envía al centro respiratorio que activa los órganos efectores: el diafragma y otros músculos de la respiración. Un mayor nivel de dióxido de carbono en la sangre, o un menor nivel de oxígeno, dará lugar a un patrón de respiración más profundo y a un aumento de la frecuencia respiratoria para devolver los gases de la sangre al equilibrio.
Un nivel demasiado bajo de dióxido de carbono y, en menor medida, un exceso de oxígeno en la sangre pueden detener temporalmente la respiración, una condición conocida como apnea, que los apneístas utilizan para prolongar el tiempo que pueden permanecer bajo el agua.
La presión parcial del dióxido de carbono es un factor más decisivo en el control del pH. Sin embargo, a gran altura (por encima de los 2.500 m) la monitorización de la presión parcial de oxígeno tiene prioridad, y la hiperventilación mantiene el nivel de oxígeno constante. Con el nivel más bajo de dióxido de carbono, para mantener el pH en 7,4 los riñones secretan iones de hidrógeno en la sangre y excretan bicarbonato en la orina. Esto es importante en la aclimatación a la altura.
Contenido de oxígeno en sangreEditar
Los riñones miden el contenido de oxígeno en lugar de la presión parcial de oxígeno en la sangre arterial. Cuando el contenido de oxígeno en la sangre es crónicamente bajo, las células sensibles al oxígeno secretan eritropoyetina (EPO) en la sangre. El tejido efector es la médula ósea roja, que produce glóbulos rojos (eritrocitos). El aumento de los glóbulos rojos conduce a un aumento del hematocrito en la sangre, y el consiguiente aumento de la hemoglobina que aumenta la capacidad de transporte de oxígeno. Este es el mecanismo por el que los habitantes de grandes altitudes tienen hematocritos más altos que los residentes a nivel del mar, y también por el que las personas con insuficiencia pulmonar o con derivaciones de derecha a izquierda en el corazón (a través de las cuales la sangre venosa evita los pulmones y pasa directamente a la circulación sistémica) tienen hematocritos igualmente altos.
Independientemente de la presión parcial de oxígeno en la sangre, la cantidad de oxígeno que puede transportarse, depende del contenido de hemoglobina. La presión parcial de oxígeno puede ser suficiente, por ejemplo, en la anemia, pero el contenido de hemoglobina será insuficiente y, posteriormente, también lo será el contenido de oxígeno. Con un aporte suficiente de hierro, vitamina B12 y ácido fólico, la EPO puede estimular la producción de glóbulos rojos, y el contenido de hemoglobina y oxígeno volver a la normalidad.
Presión arterialEditar
El cerebro puede regular el flujo sanguíneo en un rango de valores de presión arterial mediante la vasoconstricción y la vasodilatación de las arterias.
Los receptores de alta presión denominados barorreceptores en las paredes del arco aórtico y del seno carotídeo (al principio de la arteria carótida interna) controlan la presión arterial. El aumento de la presión se detecta cuando las paredes de las arterias se estiran debido a un aumento del volumen de sangre. Esto hace que las células del músculo cardíaco segreguen en la sangre la hormona péptido natriurético auricular (PNA). Esta hormona actúa sobre los riñones para inhibir la secreción de renina y aldosterona, lo que provoca la liberación de sodio y agua en la orina, reduciendo así el volumen sanguíneo. Desde aquí se estimulan los nervios motores del sistema nervioso autónomo para influir en la actividad del corazón y de las arterias de menor diámetro, llamadas arteriolas. Las arteriolas son los principales vasos de resistencia del árbol arterial, y pequeños cambios de diámetro provocan grandes cambios en la resistencia al flujo a través de ellas. Cuando la presión arterial se eleva, las arteriolas se estimulan para dilatarse, facilitando la salida de la sangre de las arterias, lo que las desinfla y hace que la presión arterial descienda, volviendo a la normalidad. Al mismo tiempo, el corazón es estimulado a través de los nervios parasimpáticos colinérgicos para que lata más despacio (lo que se denomina bradicardia), asegurando que la entrada de sangre en las arterias se reduzca, sumándose así a la reducción de la presión, y a la corrección del error original.
La baja presión en las arterias, provoca el reflejo contrario de constricción de las arteriolas, y una aceleración del ritmo cardíaco (lo que se denomina taquicardia). Si el descenso de la presión arterial es muy rápido o excesivo, la médula oblonga estimula la médula suprarrenal, a través de los nervios simpáticos «preganglionares», para que segregue epinefrina (adrenalina) en la sangre. Esta hormona potencia la taquicardia y provoca una fuerte vasoconstricción de las arteriolas de todos los órganos del cuerpo, excepto los esenciales (especialmente el corazón, los pulmones y el cerebro). Estas reacciones suelen corregir la presión arterial baja (hipotensión) de forma muy eficaz.
Niveles de calcioEditar

Los órganos efectores del primer mecanismo homeostático son los huesos, el riñón y, a través de una hormona liberada en la sangre por el riñón en respuesta a niveles elevados de PTH en la sangre, el duodeno y el yeyuno. La hormona paratiroidea (en altas concentraciones en la sangre) provoca la resorción ósea, liberando calcio en el plasma. Se trata de una acción muy rápida que puede corregir una hipocalcemia amenazante en cuestión de minutos. Las concentraciones elevadas de PTH provocan la excreción de iones de fosfato a través de la orina. Dado que los fosfatos se combinan con los iones de calcio para formar sales insolubles (véase también mineral óseo), una disminución del nivel de fosfatos en la sangre, libera iones de calcio libres en el pool de calcio ionizado del plasma. La PTH tiene una segunda acción en los riñones. Estimula la fabricación y liberación, por parte de los riñones, de calcitriol en la sangre. Esta hormona esteroidea actúa sobre las células epiteliales de la parte superior del intestino delgado, aumentando su capacidad para absorber el calcio del contenido intestinal en la sangre.
El segundo mecanismo homeostático, con sus sensores en la glándula tiroides, libera calcitonina en la sangre cuando el calcio ionizado de la sangre aumenta. Esta hormona actúa principalmente sobre el hueso, provocando la rápida extracción de calcio de la sangre y depositándolo, en forma insoluble, en los huesos.
Los dos mecanismos homeostáticos que actúan a través de la PTH, por un lado, y de la calcitonina, por otro, pueden corregir muy rápidamente cualquier error inminente en el nivel de calcio ionizado en plasma, bien extrayendo calcio de la sangre y depositándolo en el esqueleto, bien extrayendo calcio de éste. El esqueleto actúa como un almacén de calcio extremadamente grande (alrededor de 1 kg) en comparación con el almacén de calcio del plasma (alrededor de 180 mg). La regulación a largo plazo se produce a través de la absorción o la pérdida de calcio desde el intestino.
Otro ejemplo son los endocannabinoides más bien caracterizados como la anandamida (N-arachidonoylethanolamide; AEA) y el 2-arachidonoylglycerol (2-AG), cuya síntesis se produce a través de la acción de una serie de enzimas intracelulares que se activan en respuesta a un aumento de los niveles de calcio intracelular para introducir la homeostasis y la prevención del desarrollo de tumores a través de putativos mecanismos de protección que impiden el crecimiento y la migración celular mediante la activación de los receptores CB1 y/o CB2 y adyacentes.
Concentración de sodioEditar
El mecanismo homeostático que controla la concentración de sodio en plasma es bastante más complejo que la mayoría de los otros mecanismos homeostáticos descritos en esta página.
El sensor está situado en el aparato yuxtaglomerular de los riñones, que percibe la concentración de sodio en plasma de una manera sorprendentemente indirecta. En lugar de medirla directamente en la sangre que pasa por las células yuxtaglomerulares, estas células responden a la concentración de sodio en el líquido tubular renal después de que éste haya sufrido una cierta modificación en el túbulo contorneado proximal y en el asa de Henle. Estas células también responden a la tasa de flujo sanguíneo a través del aparato yuxtaglomerular, que, en circunstancias normales, es directamente proporcional a la presión sanguínea arterial, lo que convierte a este tejido en un sensor auxiliar de la presión sanguínea arterial.
En respuesta a una disminución de la concentración de sodio en plasma, o a un descenso de la presión sanguínea arterial, las células yuxtaglomerulares liberan renina en la sangre. La renina es una enzima que escinde un decapéptido (una cadena proteica corta, de 10 aminoácidos) de una α-2-globulina plasmática llamada angiotensinógeno. Este decapéptido se conoce como angiotensina I. No tiene actividad biológica conocida. Sin embargo, cuando la sangre circula por los pulmones, una enzima endotelial de los capilares pulmonares llamada enzima convertidora de angiotensina (ECA) escinde otros dos aminoácidos de la angiotensina I para formar un octapéptido conocido como angiotensina II. La angiotensina II es una hormona que actúa sobre la corteza suprarrenal, provocando la liberación en la sangre de la hormona esteroidea aldosterona. La angiotensina II también actúa sobre el músculo liso de las paredes de las arteriolas haciendo que estos vasos de pequeño diámetro se contraigan, restringiendo así la salida de sangre del árbol arterial, lo que provoca un aumento de la presión arterial. Esto, por lo tanto, refuerza las medidas descritas anteriormente (bajo el título de «Presión arterial»), que defienden la presión arterial contra los cambios, especialmente la hipotensión.
La aldosterona estimulada por la angiotensina II liberada por la zona glomerulosa de las glándulas suprarrenales tiene un efecto sobre todo las células epiteliales de los túbulos contorneados distales y los conductos colectores de los riñones. Aquí provoca la reabsorción de iones de sodio del líquido tubular renal, a cambio de iones de potasio que se secretan del plasma sanguíneo al líquido tubular para salir del organismo a través de la orina. La reabsorción de iones de sodio del líquido tubular renal detiene las pérdidas adicionales de iones de sodio del organismo y, por lo tanto, evita el empeoramiento de la hiponatremia. La hiponatremia sólo puede corregirse mediante el consumo de sal en la dieta. Sin embargo, no se sabe con certeza si la hiponatremia puede desencadenar un «hambre de sal», ni por qué mecanismo podría producirse.
Cuando la concentración plasmática de iones de sodio es superior a la normal (hipernatremia), se detiene la liberación de renina del aparato yuxtaglomerular, cesando la producción de angiotensina II, y su consiguiente liberación de aldosterona en la sangre. Los riñones responden excretando iones de sodio en la orina, normalizando así la concentración plasmática de iones de sodio. Los bajos niveles de angiotensina II en la sangre reducen la presión arterial como respuesta concomitante inevitable.
La reabsorción de iones de sodio del líquido tubular como resultado de los altos niveles de aldosterona en la sangre no provoca, por sí misma, que el agua tubular renal vuelva a la sangre desde los túbulos contorneados distales o los conductos colectores. Esto se debe a que el sodio se reabsorbe a cambio de potasio y, por tanto, sólo provoca un modesto cambio en el gradiente osmótico entre la sangre y el líquido tubular. Además, el epitelio de los túbulos contorneados distales y los conductos colectores es impermeable al agua en ausencia de la hormona antidiurética (ADH) en la sangre. La ADH forma parte del control del equilibrio de líquidos. Sus niveles en la sangre varían con la osmolalidad del plasma, que se mide en el hipotálamo del cerebro. La acción de la aldosterona en los túbulos renales impide la pérdida de sodio hacia el líquido extracelular (LEC). Por tanto, no se produce ningún cambio en la osmolalidad del LEC y, por lo tanto, tampoco en la concentración de ADH del plasma. Sin embargo, los niveles bajos de aldosterona provocan una pérdida de iones de sodio del LEC, lo que podría provocar un cambio en la osmolalidad extracelular y, por tanto, de los niveles de ADH en la sangre.
Concentración de potasioEditar
Las altas concentraciones de potasio en el plasma provocan la despolarización de las membranas de las células de la zona glomerulosa en la capa externa de la corteza suprarrenal. Esto provoca la liberación de aldosterona en la sangre.
La aldosterona actúa principalmente en los túbulos contorneados distales y en los conductos colectores de los riñones, estimulando la excreción de iones de potasio en la orina. Sin embargo, lo hace activando las bombas basolaterales de Na+/K+ de las células epiteliales tubulares. Estos intercambiadores de sodio/potasio bombean tres iones de sodio fuera de la célula hacia el líquido intersticial y dos iones de potasio hacia la célula desde el líquido intersticial. Esto crea un gradiente de concentración iónica que da lugar a la reabsorción de iones de sodio (Na+) del líquido tubular a la sangre, y a la secreción de iones de potasio (K+) de la sangre a la orina (lumen del conducto colector).
Equilibrio de líquidosEditar
La cantidad total de agua en el cuerpo debe mantenerse en equilibrio. El equilibrio de fluidos implica mantener el volumen de fluidos estabilizado, y también mantener estables los niveles de electrolitos en el líquido extracelular. El equilibrio de los fluidos se mantiene mediante el proceso de osmorregulación y mediante el comportamiento. La presión osmótica es detectada por osmorreceptores en el núcleo preóptico medio del hipotálamo. La medición de la osmolalidad del plasma para dar una indicación del contenido de agua del cuerpo, se basa en el hecho de que las pérdidas de agua del cuerpo, (a través de la pérdida inevitable de agua a través de la piel que no es totalmente impermeable y por lo tanto siempre ligeramente húmeda, el vapor de agua en el aire exhalado, la sudoración, los vómitos, las heces normales y especialmente la diarrea) son todos hipotónicos, lo que significa que son menos salados que los fluidos corporales (comparar, por ejemplo, el sabor de la saliva con el de las lágrimas. Esta última tiene casi el mismo contenido en sal que el líquido extracelular, mientras que la primera es hipotónica con respecto al plasma. La saliva no tiene un sabor salado, mientras que las lágrimas son decididamente saladas). Por lo tanto, casi todas las pérdidas normales y anormales de agua corporal hacen que el líquido extracelular se vuelva hipertónico. Por el contrario, la ingesta excesiva de líquidos diluye el líquido extracelular haciendo que el hipotálamo registre condiciones de hiponatremia hipotónica.
Cuando el hipotálamo detecta un entorno extracelular hipertónico, provoca la secreción de una hormona antidiurética (ADH) llamada vasopresina que actúa sobre el órgano efector, que en este caso es el riñón. El efecto de la vasopresina en los túbulos renales es reabsorber el agua de los túbulos contorneados distales y de los conductos colectores, evitando así que se agrave la pérdida de agua por la orina. Al mismo tiempo, el hipotálamo estimula el centro de la sed cercano, provocando un impulso casi irresistible (si la hipertonicidad es lo suficientemente grave) de beber agua. El cese del flujo de orina impide que la hipovolemia y la hipertonicidad empeoren; el hecho de beber agua corrige el defecto.
La hipoosmolalidad da lugar a niveles de ADH en plasma muy bajos. Esto provoca la inhibición de la reabsorción de agua de los túbulos renales, lo que hace que se excreten volúmenes elevados de orina muy diluida, deshaciéndose así del exceso de agua en el organismo.
La pérdida de agua por vía urinaria, cuando el homeostato del agua corporal está intacto, es una pérdida de agua compensatoria, que corrige cualquier exceso de agua en el organismo. Sin embargo, dado que los riñones no pueden generar agua, el reflejo de la sed es el importantísimo segundo mecanismo efector del homeostato del agua corporal, corrigiendo cualquier déficit de agua en el cuerpo.
Hemostasis de la sangre
El pH plasmático puede ser alterado por cambios respiratorios en la presión parcial de dióxido de carbono; o alterado por cambios metabólicos en la relación de ácido carbónico a ion bicarbonato. El sistema de amortiguación del bicarbonato regula la relación entre el ácido carbónico y el bicarbonato para que sea igual a 1:20, en cuya relación el pH de la sangre es de 7,4 (como se explica en la ecuación de Henderson-Hasselbalch). Un cambio en el pH plasmático produce un desequilibrio ácido-base.En la homeostasis ácido-base hay dos mecanismos que pueden ayudar a regular el pH. La compensación respiratoria, un mecanismo del centro respiratorio, ajusta la presión parcial de dióxido de carbono cambiando el ritmo y la profundidad de la respiración, para devolver el pH a la normalidad. La presión parcial de dióxido de carbono también determina la concentración de ácido carbónico, y el sistema tampón de bicarbonato también puede entrar en juego. La compensación renal puede ayudar al sistema tampón de bicarbonato.El sensor de la concentración plasmática de bicarbonato no se conoce con certeza. Es muy probable que las células tubulares renales de los túbulos contorneados distales sean a su vez sensibles al pH del plasma. El metabolismo de estas células produce dióxido de carbono, que se convierte rápidamente en hidrógeno y bicarbonato mediante la acción de la anhidrasa carbónica. Cuando el pH del ECF desciende (volviéndose más ácido), las células tubulares renales excretan iones de hidrógeno en el líquido tubular para salir del cuerpo a través de la orina. Al mismo tiempo, se secretan iones de bicarbonato en la sangre, lo que disminuye el ácido carbónico y, en consecuencia, eleva el pH plasmático. Lo contrario ocurre cuando el pH del plasma se eleva por encima de lo normal: los iones de bicarbonato se excretan en la orina, y los iones de hidrógeno se liberan en el plasma.
Cuando los iones de hidrógeno se excretan en la orina, y el bicarbonato en la sangre, este último se combina con el exceso de iones de hidrógeno en el plasma que estimuló a los riñones a realizar esta operación. La reacción resultante en el plasma es la formación de ácido carbónico que está en equilibrio con la presión parcial plasmática de dióxido de carbono. Esto se regula estrechamente para garantizar que no haya una acumulación excesiva de ácido carbónico o bicarbonato. Por lo tanto, el efecto global es que los iones de hidrógeno se pierden en la orina cuando el pH del plasma desciende. El aumento concomitante del bicarbonato plasmático absorbe el aumento de los iones de hidrógeno (causado por el descenso del pH plasmático) y el exceso de ácido carbónico resultante se elimina en los pulmones en forma de dióxido de carbono. Esto restablece la relación normal entre el bicarbonato y la presión parcial de dióxido de carbono y, por tanto, el pH plasmático.Lo contrario ocurre cuando un pH plasmático elevado estimula a los riñones a secretar iones de hidrógeno en la sangre y a excretar bicarbonato en la orina. Los iones de hidrógeno se combinan con el exceso de iones de bicarbonato en el plasma, formando de nuevo un exceso de ácido carbónico que puede ser exhalado, en forma de dióxido de carbono, en los pulmones, manteniendo constante la concentración de iones de bicarbonato en el plasma, la presión parcial de dióxido de carbono y, por tanto, el pH plasmático.
Líquido cefalorraquídeoEditar
El líquido cefalorraquídeo (LCR) permite regular la distribución de sustancias entre las células del cerebro, y los factores neuroendocrinos, a los que ligeras alteraciones pueden causar problemas o daños en el sistema nervioso. Por ejemplo, una concentración elevada de glicina altera el control de la temperatura y la presión arterial, y un pH elevado del LCR provoca mareos y síncopes.
NeurotransmisiónEditar
Las neuronas inhibidoras del sistema nervioso central desempeñan un papel homeostático en el equilibrio de la actividad neuronal entre la excitación y la inhibición. Las neuronas inhibitorias, mediante el GABA, realizan cambios compensatorios en las redes neuronales impidiendo los niveles desbocados de excitación. Un desequilibrio entre la excitación y la inhibición se ve implicado en una serie de trastornos neuropsiquiátricos.
Sistema neuroendocrinoEditar
El sistema neuroendocrino es el mecanismo por el que el hipotálamo mantiene la homeostasis, regulando el metabolismo, la reproducción, la conducta de comer y beber, la utilización de la energía, la osmolaridad y la presión arterial.
La regulación del metabolismo, se lleva a cabo mediante interconexiones hipotalámicas con otras glándulas. Tres glándulas endocrinas del eje hipotálamo-hipofisario-gonadal (eje HPG) suelen trabajar juntas y tienen importantes funciones reguladoras. Otros dos ejes endocrinos reguladores son el eje hipotálamo-hipófisis-suprarrenal (eje HPA) y el eje hipotálamo-hipófisis-tiroides (eje HPT).
El hígado también tiene muchas funciones reguladoras del metabolismo. Una función importante es la producción y el control de los ácidos biliares. Demasiado ácido biliar puede ser tóxico para las células y su síntesis puede ser inhibida por la activación de FXR un receptor nuclear.
Regulación génicaEditar
A nivel celular, la homeostasis se lleva a cabo por varios mecanismos, entre ellos la regulación transcripcional que puede alterar la actividad de los genes en respuesta a los cambios.
Equilibrio energéticoEditar
La cantidad de energía ingerida a través de la alimentación tiene que coincidir con la cantidad de energía utilizada. Para lograr la homeostasis energética el apetito se regula mediante dos hormonas, la grehlin y la leptina. La grehlin estimula el hambre y la ingesta de alimentos y la leptina actúa para señalar la saciedad (plenitud).
Una revisión de 2019 sobre las intervenciones de cambio de peso, incluyendo las dietas, el ejercicio y la sobrealimentación, descubrió que la homeostasis del peso corporal no podía corregir con precisión los «errores energéticos», la pérdida o el aumento de calorías, a corto plazo.