Dale Vogt, Helen A. Swartz and John Massey
Department of Animal Sciences
Vari schemi di accoppiamento degli animali sono classificati sotto due grandi categorie – inbreeding e outbreeding. La classificazione dipende dalla vicinanza della relazione biologica tra i compagni. All’interno di ogni categoria, esiste un’ampia variazione nell’intensità di questa relazione. Una linea molto sottile separa le due categorie. L’accoppiamento di animali strettamente imparentati (per esempio, genitore e prole, fratello e sorella completi o mezzo fratello e sorella) è inbreeding. Con animali meno strettamente imparentati (cugini di primo grado, cugini di secondo grado), le persone non sono d’accordo su dove tracciare la linea tra inbreeding e outbreeding.
Tecnicamente, l’inbreeding è definito come l’accoppiamento di animali più strettamente imparentati del rapporto medio all’interno della razza o della popolazione interessata. Gli accoppiamenti tra animali meno strettamente imparentati di questo, quindi, costituirebbero outbreeding. Questi due sistemi di accoppiamento, con diverse intensità in ciascuno, sono descritti nella tabella 1. Gli accoppiamenti indicati all’interno della categoria inbreeding sono autoesplicativi; quelli all’interno della categoria outbreeding sono definiti nel glossario.
Tabella 1
Gradi di inbreeding e outbreeding disposti secondo la relazione biologica tra gli accoppiamenti indicati. (Leggendo dall’alto verso il basso, la relazione biologica tra compagni diminuisce costantemente.)
| Inbreeding o outbreeding | Coefficiente di relazione tra compagni | Rapporto biologico tra compagni | Rapporto biologico relazione tra compagni |
|---|---|---|---|
| Inbreeding | 50 per cento | Padre × prole; sorelle complete | |
| Inbreeding | 25 per cento | Mezze sorelle; doppie cugine di primo grado; zia × nipote; zio × nipote | |
| Inbreeding | 12-1/2 per cento | Cugini di primo grado | |
| Inbreeding | 6-1/4 per cento | Cugini di secondo grado | |
| Inbreeding | ? | Linebreeding1 | |
| Inbreeding/outbreeding | 0 per cento | Abbinamento casuale all’interno della razza o della popolazione2 | |
| Outbreeding | 0 per cento | Outcrossing | |
| Outbreeding | 0 per cento | Incrocio di razza | |
| Outbreeding | 0 per cento | Incrocio di specie | |
| Outbreeding | 0 per cento | Incrocio di genere |
1In un programma di linebreeding, il coefficiente di relazione tra compagni è solitamente basso; tuttavia, può essere molto variabile.
2L’accoppiamento casuale all’interno di una razza o di una popolazione significa che i compagni sono scelti per caso. Bisogna capire che in questa circostanza è possibile che si verifichino sia inbreeding che outbreeding.
Relazioni biologiche tra animali
Gli individui sono considerati biologicamente correlati quando hanno uno o più antenati comuni. Per scopi pratici, se due individui non hanno un antenato comune nelle ultime cinque o sei generazioni, sono considerati non imparentati.
La relazione biologica è importante nell’allevamento degli animali perché più stretta è la relazione, più alta è la percentuale di geni simili che i due individui portano. La vicinanza di parentela è determinata da tre fattori:
- Quanto indietro nei pedigree dei due animali appare l’antenato comune
- Quanti antenati comuni hanno
- Quanto frequentemente appaiono gli antenati comuni. È anche influenzato dall’eventuale inbreeding dell’antenato o degli antenati comuni
Misura del grado di relazione biologica
Il coefficiente di relazione è un singolo valore numerico che considera tutti i fattori sopra menzionati. È una misura del grado in cui i genotipi (costituzioni genetiche) dei due animali sono simili. Si stima con l’espressione:
RBC = sigma ÷ Radice quadrata di (1 + FB)(1 + FC)
Equazione 1 dove:
RBC = il coefficiente di relazione tra gli animali B e C che vogliamo misurare.
sigma = il simbolo greco che significa “aggiungere”.
(1/2) = la frazione del materiale genetico di un individuo che viene trasmesso alla sua progenie. È usato nel calcolo del coefficiente di parentela perché rappresenta la probabilità che, in una qualsiasi generazione, un gene identico di una data coppia di geni sia trasmesso a ciascuna di due progenie particolari. È anche la probabilità che un gene diverso da una data coppia di geni sia trasmesso alle due progenie.
n = il numero di generazioni tra l’animale B e l’antenato comune.
n’ = il numero di generazioni tra l’animale C e l’antenato comune
FA, FB, FC = coefficienti di inbreeding dell’antenato comune e degli animali B e C, rispettivamente.
Se nessuno degli animali è consanguineo, il coefficiente di parentela è stimato come:
RBC = sigma
Equazione 2
L’uso di questa espressione può essere dimostrato con il pedigree campione full-sib e il diagramma a freccia (Figura 1). In questo esempio, supponiamo che né il padre né la madre siano consanguinei. Il diagramma a freccia sulla destra mostra i percorsi del flusso genico da ciascuno degli antenati comuni (D ed E) agli animali di cui stiamo misurando il coefficiente di parentela (B e C).
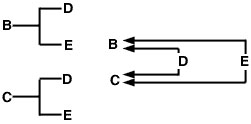
Figura 1
Il problema ora è di tracciare tutti i possibili percorsi dall’animale B all’animale C che passano attraverso un antenato comune. In questo caso, ci sono due percorsi di questo tipo (figura 2).
Figura 2
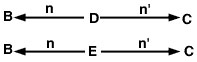
Equazione 3
Siccome in questo esempio abbiamo ipotizzato l’assenza di inbreeding, il coefficiente di relazione tra gli animali B e C viene stimato come:
RBC = sigma = (1/2)1 + 1 + (1/2)1 + 1 = (1/2)2 + (1/2)2 = 0.50
Utilità delle informazioni sul coefficiente di rapporto
Un produttore di bestiame troverebbe le informazioni sul coefficiente di rapporto preziose in diverse situazioni. Potrebbe, per esempio, voler vendere un animale imparentato con uno che precedentemente ha venduto ad un prezzo elevato. Più alto è il coefficiente di relazione tra i due, migliore è il suo utilizzo come punto di vendita. Oppure, potrebbe voler acquistare uno dei due tori imparentati e uno potrebbe costare più di quanto voglia pagare. Se il coefficiente di relazione tra i due tori è alto, potrebbe trovarsi bene sia con il toro di prezzo inferiore che con quello più costoso.
Un uso pratico del coefficiente di relazione è la stima del valore di performance di un animale non testato. Per stimare il valore, dobbiamo conoscere il valore di performance di un animale correlato, il coefficiente di relazione tra gli animali testati e non testati, e il valore di performance medio della razza, della mandria o del gruppo a cui appartengono gli animali testati e non testati.
Come esempio, consideriamo una mandria con un guadagno medio giornaliero (ADG) di 2,25 libbre al giorno. Supponiamo inoltre che un padre di questa mandria abbia un ADG di 3,50 libbre al giorno, mentre un fratellastro più giovane non è ancora stato valutato per l’ADG. Supponendo che non ci sia consanguineità, il coefficiente di relazione tra i fratellastri è 0,25. La migliore stima dell’ADG dell’animale non testato è che si discosterà dalla media della mandria del 25 per cento quanto il valore delle prestazioni del fratellastro testato. Usando queste cifre, il più probabile valore ADG dell’animale non testato è 2,25 + (0,25) × (3,50 – 2,25) o 2,56 libbre al giorno.
Misura del grado di inbreeding
Quando calcoliamo un coefficiente di inbreeding, stiamo cercando di misurare la probabile riduzione percentuale della frequenza di accoppiamento di geni dissimili (riduzione dell’eterozigosi). Questa riduzione è relativa a una popolazione di base. La popolazione di base di solito è la razza interessata ad una data a cui i pedigree sono tracciati. Si presume che gli animali di questa popolazione di base non siano incrociati. Questo non significa che questi animali della popolazione di base avevano geni dissimili in ogni coppia. Non c’è modo per noi di sapere quante delle loro coppie di geni consistevano in geni simili o dissimili. Il coefficiente di inbreeding che viene calcolato è semplicemente relativo a quella base e riflette la probabile riduzione percentuale di quante coppie di geni dissimili avevano in media gli animali della popolazione base.
L’espressione generale per determinare il coefficiente di inbreeding è:
FX = sigma
Equazione 4 dove:
FX = il coefficiente di inbreeding dell’animale X.
sigma = il simbolo greco che significa “aggiungere”.
(1/2) = la frazione del materiale genetico di un individuo che viene trasmesso alla sua prole. Si usa nel calcolo del coefficiente di parentela perché rappresenta la probabilità che, in una qualsiasi generazione, un gene identico di una data coppia di geni sia trasmesso a ciascuna di due progenie particolari. È anche la probabilità che un gene diverso da una data coppia di geni sia trasmesso alle due progenie.
n = il numero di generazioni tra l’animale B e l’antenato comune.
n’ = il numero di generazioni tra l’animale C e l’antenato comune.
+1 = si aggiunge a n e n’ per tenere conto della generazione supplementare tra l’animale × e i suoi genitori.
FA = il coefficiente di consanguineità dell’antenato comune.
Se nessuno dei due genitori è consanguineo, ma se sono imparentati, il coefficiente di consanguineità della loro prole è la metà del loro coefficiente di parentela: 1/2 RBC Questo può essere dimostrato usando un accoppiamento full-sib per facilitare il confronto con il coefficiente di parentela full-sib calcolato in precedenza. In questo caso, il pedigree dell’animale X e il diagramma delle frecce saranno i seguenti:
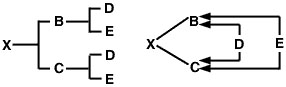
Figura 3
Il problema ora è di tracciare tutti i possibili percorsi dal padre (B) alla madre (C) attraverso ogni antenato comune. Come per il problema del coefficiente di parentela, ci sono due percorsi:
![]() Figura 4
Figura 4
Poiché abbiamo assunto che nessuno dei due genitori sia consanguineo, il coefficiente di inbreeding dell’animale × è stimato come:
FX = sigma = (1/2)1 + 1 + 1 + 1 + (1/2)1 + 1 + 1 = (1/2)3 + (1/2)3 = 0.25
Equazione 5
Questo è la metà del coefficiente di relazione tra i full-sibs quando non c’è inbreeding.
Conseguenze genetiche dell’inbreeding
La conseguenza genetica di base dell’inbreeding è di promuovere ciò che è tecnicamente noto come omozigosi. Questo significa che c’è un aumento della frequenza di accoppiamento di geni simili. Accompagnando questo aumento, ci deve essere una diminuzione della frequenza di accoppiamento dei geni dissimili. Questo è chiamato una diminuzione dell’eterozigosi. Questi eventi simultanei sono le ragioni alla base degli effetti generali sulla performance che osserviamo con l’inbreeding.
Motivi per l’inbreeding
Lo sviluppo di linee di bestiame domestico altamente produttive è possibile. Fino ad oggi, tuttavia, tali tentativi hanno avuto poco successo apparente. Anche se occasionalmente si producono animali ad alte prestazioni, l’inbreeding generalmente si traduce in una riduzione complessiva delle prestazioni. Questa riduzione si manifesta in molti modi. Gli effetti più ovvi dell’inbreeding sono una minore efficienza riproduttiva, compresi tassi di mortalità più elevati, tassi di crescita più bassi e una maggiore frequenza di anomalie ereditarie. Questo è stato dimostrato da numerosi studi su bovini, cavalli, pecore, suini e animali da laboratorio.
L’entità di questa diminuzione della performance, in generale, è proporzionale al grado di inbreeding. Maggiore è il grado di inbreeding, maggiore è la riduzione della performance. La riduzione effettiva delle prestazioni non è la stessa in tutte le specie o in tutti i tratti. Alcune caratteristiche (come la qualità della carne) sono appena influenzate dall’inbreeding; altre (come l’efficienza riproduttiva) sono fortemente influenzate dall’inbreeding. Non possiamo, quindi, fare un’affermazione generalizzata sulla quantità di riduzione della “performance” che risulterebbe da una specifica quantità di inbreeding e aspettarci che sia applicabile in un’ampia varietà di situazioni.
È possibile, tuttavia, prevedere l’entità dell’effetto dell’inbreeding su tratti specifici. Tali previsioni sono basate su risultati effettivamente ottenuti in condizioni sperimentali in cui sono stati raggiunti vari livelli di inbreeding. In una ricerca sui suini condotta presso il Midwest Regional Swine Breeding Laboratory, Dickerson e altri (1954) sottolineano che per ogni aumento del 10% di inbreeding (dei suini nella cucciolata), c’è una diminuzione di 0.20, 0.35, 0.38, e 0.44 suini per cucciolata alla nascita, 21 giorni, 56 giorni, e 154 giorni, rispettivamente. Possiamo usare queste cifre per fornire stime delle diminuzioni attese nelle dimensioni della cucciolata (a età comparabili) in altre mandrie di suini.
La maggior parte degli studi sull’inbreeding suggerisce che ogni aumento successivo di unità di inbreeding si traduce in una diminuzione proporzionale della performance. Le stime degli aumenti medi della percentuale di inbreeding all’interno di una mandria chiusa possono essere fatte con l’espressione:
Avg. aumento di inbreeding = (Numero maschi + Numero femmine) ÷ (8)(Numero maschi)(Numero femmine)
In una mandria chiusa di bovini in cui 100 femmine e quattro maschi sono stati usati in ogni generazione, per esempio, l’aumento medio per generazione di inbreeding sarebbe (4 + 100) ÷ (8)(4)(100) = 0.0325. Su base annua, assumendo un intervallo di generazione di cinque anni, ciò equivarrebbe ad un aumento medio annuo di inbreeding di 0,0065 o 0,65%.
Nonostante i risultati generalmente scarsi ottenuti con l’inbreeding, esso è uno strumento molto utile nell’allevamento degli animali. L’inbreeding è essenziale per lo sviluppo di animali prepotenti – animali che “imprimono” uniformemente le loro caratteristiche sulla loro progenie. Poiché l’inbreeding causa un aumento della proporzione di geni simili (buoni o cattivi, recessivi o dominanti), le cellule riproduttive dell’animale inbreeding saranno più uniformi nel loro corredo genetico. Quando questa uniformità coinvolge un numero relativamente grande di geni dominanti, la progenie di quell’individuo mostrerà uniformemente le caratteristiche dominanti di quel genitore.
L’inbreeding può anche essere usato per scoprire i geni che producono anormalità o morte – geni che, negli allevamenti outbred, sono generalmente presenti in basse frequenze. Questi geni dannosi sono quasi sempre recessivi nella loro natura genetica e i loro effetti sono nascosti o mascherati dalle loro controparti dominanti (alleli). Ad eccezione dei tratti legati al sesso, i geni recessivi non sono espressi se portati singolarmente. Perché i loro effetti si manifestino, devono essere presenti in doppio. La probabilità che siano presenti in duplicato aumenta con l’inbreeding, perché l’inbreeding aumenta la proporzione di geni simili (sia buoni che cattivi) nella popolazione inbred. Una volta scoperti gli effetti di questi geni, l’allevatore può eliminarli dalla sua mandria. Abbatterebbe la progenie che mostra l’effetto indesiderato di questi geni recessivi e abbatterebbe anche i genitori che sono portatori dei geni indesiderati. Inoltre, due terzi della progenie “normale” di questi genitori portatori sono a loro volta portatori di questi stessi geni indesiderabili. In assenza di test d’allevamento per distinguere i portatori dai non portatori, sarebbe anche necessario abbattere tutta la progenie “normale” dei genitori portatori.
Gli allevatori possono usare un test d’inbreeding per identificare i portatori di geni autosomici recessivi dannosi (come quelli responsabili del nanismo di Snorter nei bovini, dell’iperostosi nei suini, o del criptorchidismo nelle pecore). Un test di inbreeding controlla solo i geni recessivi che l’animale testato (di solito il maschio) porta. L’esempio seguente e i numeri indicati sono pertinenti al bestiame, dove una prole per gestazione è usuale. Se vogliamo essere sicuri al livello di probabilità 0,01 che un toro sia privo di geni recessivi autosomici dannosi, dovremmo accoppiarlo con almeno 35 delle sue figlie. Viene accoppiato con più di 35 delle sue figlie perché solo la metà di loro dovrebbe essere portatrice di qualsiasi gene recessivo dannoso di cui è portatore il loro padre. Abbiamo bisogno della produzione di 35 vitelli normali senza un solo vitello anormale per dimostrare che il toro è esente (al livello di probabilità 0,01) da qualsiasi gene recessivo autosomico dannoso.
I test nelle pecore e nei maiali richiederebbero l’accoppiamento con meno figlie. Il numero effettivo dipenderebbe dal numero medio di progenie prodotte per gestazione in ogni specie. Usando il livello di probabilità 0,01 e assumendo che la dimensione media della cucciolata sia 1,5 nelle pecore e 8,0 nei suini, il numero di accoppiamenti padre-figlia necessari sarebbe di circa 24+ per le pecore e 5+ per i suini.
Un altro uso importante dell’inbreeding è nello sviluppo di famiglie distinte o linee inbred. Iniziando con una popolazione genetica inizialmente diversa, l’inbreeding porta alla formazione di varie linee, ognuna delle quali differisce geneticamente dall’altra. L’inbreeding continuo all’interno di queste linee tende a cambiare la frequenza di alcuni dei geni presenti nella popolazione iniziale. Per esempio, se un gene particolare è presente solo nell’1% degli animali della popolazione iniziale, l’inbreeding e lo sviluppo di linee distinte potrebbero far sì che questo gene sia presente in tutti o quasi tutti gli animali di alcune linee e in nessuno o solo alcuni degli animali di altre linee. Le linee consanguinee sono utilizzate in vari modi, ma probabilmente sono usate soprattutto nello sviluppo di polli ibridi o di mais da semina ibrido.
Una forma generalmente blanda di inbreeding (linebreeding) è usata con successo da alcuni produttori di sementi e commerciali. Il suo obiettivo è quello di mantenere un alto grado di parentela tra gli animali della mandria e qualche antenato o antenati eccezionali. Con l’inbreeding in generale, non c’è alcun tentativo di aumentare la relazione tra la prole e un particolare antenato. In un programma di linebreeding c’è un tentativo deliberato di mantenere o aumentare la relazione tra la prole e uno specifico antenato ammirato (o antenati). Questa caratteristica distingue il linebreeding come una forma speciale di inbreeding.
Il coefficiente di inbreeding della prole prodotta in un programma di linebreeding è generalmente basso, ma dipende dal tipo di programma di allevamento seguito. La figura 5 illustra due programmi di linebreeding. Il programma d’allevamento delineato nella parte A mostra una linea di relazione diretta tra la prole “x” e l’antenato desiderato “5”. Si raggiunge solo un lieve livello di inbreeding nell’animale “x” (Fx = 0,03125). Il coefficiente di parentela ottenuto tra l’animale “x” e l’animale “5” è 0,2462. La parte B è un programma di linebreeding basato su accoppiamenti continui padre-figlia che, dopo solo due generazioni, produce un livello di inbreeding nell’animale “x” di 0,375 e un coefficiente di relazione tra gli animali “x” e “S” di 0,78.
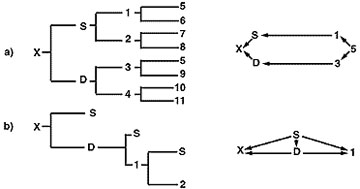 Figura 5
Figura 5
Pedigree e diagrammi di freccia per due programmi di linebreeding.
Il programma padre-figlia nella parte B concentra efficacemente i geni dell’animale “S”, ma a causa del rapido aumento dell’inbreeding della progenie prodotta in questo programma, l’allevatore corre il rischio di prestazioni molto ridotte e un’alta probabilità di difetti genetici. I programmi di linebreeding sono utilizzati al meglio nelle mandrie di purosangue e ad alte prestazioni quando un individuo geneticamente superiore in quella mandria è stato identificato e valutato dai test di progenie. Concentrare i geni di quell’individuo sarebbe poi meglio realizzato accoppiandolo con femmine non imparentate per ridurre il rischio di effetti dannosi associati a tale intenso inbreeding.
Riassunto
Inbreeding è tecnicamente definito come l’accoppiamento di animali più strettamente legati rispetto alla relazione media all’interno della razza o della popolazione interessata. Per scopi pratici, se due individui accoppiati non hanno un antenato comune entro le ultime cinque o sei generazioni, la loro progenie sarebbe considerata outbred. La principale conseguenza genetica dell’inbreeding è l’aumento della frequenza di accoppiamento di geni simili. Tutti i cambiamenti genetici e fenotipici associati alla pratica dell’inbreeding derivano da questa unica conseguenza primaria. In generale, l’inbreeding si traduce in un abbassamento generale della performance. Si riflette più ovviamente in una minore efficienza riproduttiva, compresi tassi di mortalità più alti, tassi di crescita più bassi e una maggiore frequenza di difetti ereditari. Nonostante questi effetti generalmente dannosi, l’inbreeding è uno strumento molto utile nel campo dell’allevamento animale. Permette all’allevatore di scoprire ed eliminare i geni recessivi dannosi all’interno della popolazione. È anche essenziale per lo sviluppo di animali prepotenti ed è auspicabile nello sviluppo di linee familiari distinte. Inoltre, i produttori di sementi e commerciali hanno usato con successo il linebreeding per mantenere un certo grado di relazione genetica nei loro animali con uno o più antenati eccezionali.
Glossario
I termini descrivono vari schemi di accoppiamento e la progenie risultante da specifici programmi di accoppiamento.
- Backcross
Progressione risultante dall’accoppiamento di un animale di due razze incrociate con una delle razze parentali. Per esempio, usando due razze designate come P1 e P2, la progenie backcross sarebbe prodotta dall’accoppiamento dell’animale incrociato di due razze (Pl × P2) con una delle razze parentali P1 o P2. - Crisscrossing
Un programma continuo di incrocio in cui vi è un uso alternato di maschi appartenenti a due razze. Utilizzando due razze designate come P1 e P2, un programma di incrocio, iniziando con l’incrocio di due razze (P1 × P2), inizierebbe con un backcrossing con una delle razze parentali. Le femmine risultanti da questi accoppiamenti sarebbero allevate con un maschio P2, × P2, e così via. - Incrocio
Progetto risultante dall’accoppiamento di animali outcross appartenenti a razze diverse. - Incrocio di genere
Abbinamento di animali appartenenti a generi diversi (per esempio, l’accoppiamento di bovini domestici, Bos taurus o Bos indicus, con il bisonte americano, Bison). - Classificazione
Abbinamento di maschi purosangue di una data razza con femmine non purosangue e la risultante prole femminile in generazioni successive. - Linea inbred
Linea di animali prodotta dall’accoppiamento di animali correlati. - Inbreeding
Abbinamento di animali più strettamente imparentati tra loro rispetto al rapporto medio all’interno della razza o della popolazione in questione. - Incrocio
Progressione risultante dall’accoppiamento di animali di diverse linee inbred all’interno di una razza. - Incrocio
Progressione risultante dall’accoppiamento di animali provenienti da linee consanguinee di razze diverse. - Linebreeding
Forma generalmente leggera di inbreeding in cui gli animali accoppiati sono imparentati con qualche individuo presumibilmente eccezionale. - Outbreeding
Abbinamento di animali meno strettamente imparentati tra loro rispetto al rapporto medio all’interno della razza o popolazione interessata. - Outcross
Progressione risultante dall’accoppiamento di animali non imparentati all’interno di una razza. - Incrocio di specie
Abbinamento di animali appartenenti a specie diverse (per esempio l’accoppiamento di bovini di razza europea, Bos taurus, con bovini Brahman, Bos indicus). - Incrocio a rotazione di tre razze
Un programma continuo di incrocio in cui i maschi di tre razze sono usati a rotazione. Usando tre razze designate come P1, P2 e P3, la prima generazione comporterebbe la produzione di animali di due razze incrociate, P1 x P2. Nella seconda generazione, le femmine dell’incrocio di due razze verrebbero accoppiate con maschi della terza razza, (P1 × P2) × P3; le femmine dell’incrocio di tre razze verrebbero accoppiate con maschi di una delle razze usate per produrre gli animali dell’incrocio di due razze, × P1, e così via. - Topcross
Progressione risultante dall’accoppiamento di animali appartenenti a famiglie diverse all’interno di una razza. - Topcrossbred
Progressione risultante dall’accoppiamento di maschi consanguinei con femmine non consanguinee di un’altra razza. - Topincross
Progressione risultante dall’accoppiamento di maschi consanguinei con femmine non consanguinee della stessa razza. - Incrocio di due razze
Progressione risultante dall’accoppiamento di maschi di una razza con femmine di un’altra razza.